von Dr. Heinrich Niehaus, Celle-Lachtehausen – „Das Blaue Jahrbuch“1979
Die Vererbungslehre ist eine noch relativ junge Wissenschaft. Erst nach Wiederentdeckung der Mendelgesetze durch Correns, De Vries und Tschermak hat man sich seit Beginn des 20. Jahrhunderts in allen Teilen der Welt intensiv mit der weiteren Erforschung der Erbvorgänge befasst und die Entwicklung der modernen Vererbungslehre eingeleitet.
Inzwischen ist aus der zunächst zarten Pflanze ein mächtiger Baum mit vielen Zweigen und Ästen geworden, ein umfangreiches Stoffgebiet, dessen ausführliche Darstellung viele dicke Bände füllen würde. So ist es z. B. gelungen, mit Hilfe des Elektronenmikroskops, feinsinniger Analysenmethoden und Datenverarbeitungsanlagen in die mikroskopische Welt der Erbsubstanz einzudringen und ihren atomaren Aufbau und ihre komplizierte Wirkungsweise zu entdecken.
Die auf diesem als Molekulargenetik bezeichneten Gebiet gesammelten Erkenntnisse und Erfahrungen ermöglichten dann Versuche zur gezielten Veränderung des Erbgutes. Von den Zielen solcher besonders an Bakterien, aber auch an Säugetieren vor- genommenen Experimente seien besonders die Erforschung und Bekämpfung der Krebskrankheiten genannt. Zu den weiteren Zweigen der Genetik gehören die Zytogenetik (Zellgenetik), Mutationsgenetik, Populationsgenetik, Humangenetik und andere.
Ich werde mich im Rahmen dieses Artikels darauf beschränken, auf einige mir wichtig erscheinende Grundelemente der Vererbung hinzuweisen, Gesetzmäßigkeiten bei einfachen Erbvorgängen aufzuzeigen und diese an Beispielen aus der Kaninchenzucht zu erläutern.
Die große Bedeutung, welche die Kenntnisse der Vererbungsvorgänge für eine gezielte Züchtung, speziell auch in der Rassekaninchenzucht, besitzen, dürften allen erfahrenen Kaninchenzüchtern hinlänglich bekannt sein.
Was ist Vererbung?
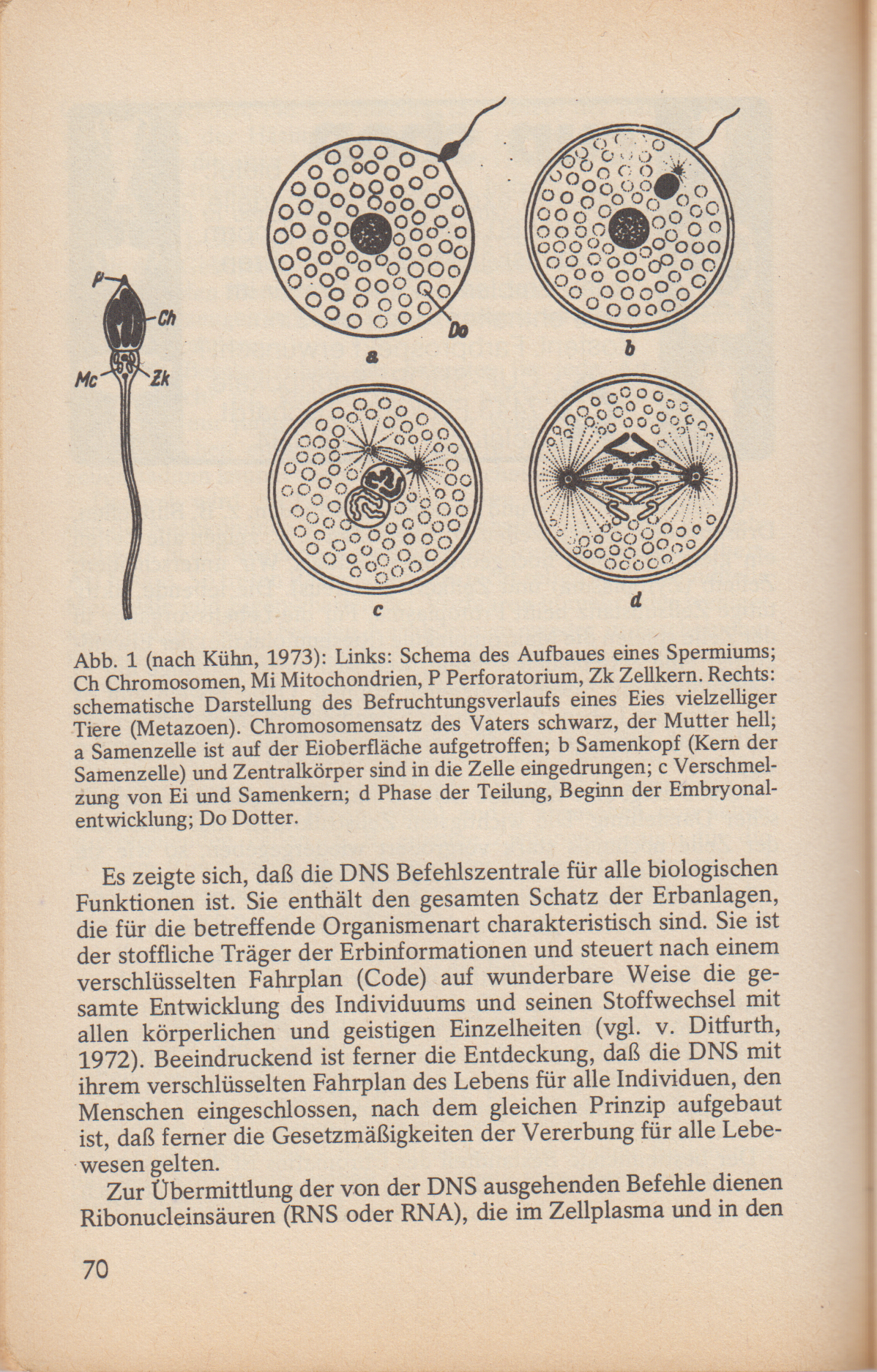
Als Vererbung bezeichnet man die Weitergabe von Erbanlagen (Gene) als Träger von Erbinformationen von den Vorfahren auf die Nachkommen. Diese Weitergabe erfolgt bei Kaninchen und anderen Tieren mit geschlechtlicher Fortpflanzung durch die Keim- oder Geschlechtszellen, die Samenzellen (Spermien) der Rammler und die Eizellen der Häsinnen. Nach einer gelungenen Paarung vereinigen sich die etwa 10-15 Eizellen der Häsin mit je einer Samenzelle des Rammlers (Abb. 1). Aus so befruchteten Eizellen entwickeln sich in der Gebärmutter (Uterus) der Häsin junge Kaninchen, die 31-32 Tage nach der Paarung den mütterlichen Körper durch den Geburtsakt verlassen, in weiteren 4-7 Monaten zu geschlechtsreifen Tieren heranwachsen, um dann selbst wieder Nachkommen erzeugen bzw. gebären zu können. So folgt eine Generation auf die andere.
Da die Beschaffenheit des Erbgutes für jede Art spezifisch (eigentümlich) ist, können sich aus befruchteten Eizellen von Kaninchen nur junge Kaninchen, aber keine Ratten, Mäuse oder Gänseblümchen entwickeln. Für andere Arten, die sich im Eizellenstadium kaum voneinander unterscheiden, gilt dasselbe.
Dadurch wird die Erhaltung der Arten gesichert und bewirkt, dass die Nachkommen ihren Vorfahren ähnlich sind. Diese Ähnlichkeit geht in der Regel noch weit über die Grundmerkmale der betr. Art hinaus. So treten Rassemerkmale, nicht selten auch individuelle Merkmale und Eigenschaften der Vorfahren in mehr oder weniger ausgeprägter Form bei den Nachkommen wieder in Erscheinung. Beispiele: Stirnfleck, Kopfform, abfallende oder abgerundete Kruppe, ruhiges oder nervöses Wesen, robuste Gesundheit oder Anfälligkeit gegen Krankheiten usw.
Trotz mancher verblüffender Ähnlichkeiten zwischen Vorfahren und Nachkommen gibt es jedoch keine völlige Gleichheit innerhalb der Arten, Rassen, Stämme und Würfe. Das gilt ohne Ausnahme für das Erscheinungsbild (Phänotypus) und abgesehen von selten vorkommenden Fällen (z. B. eineiige Zwillinge) auch für die Erbform (Genotypus). Wie es zu dieser Ungleichheit kommt und welche Bedeutung sie für die Züchtung besitzt, werden wir im Folgenden noch näher kennenlernen.
Die Zelle als Träger der Erbanlagen
Die Zelle (Cellula, kleine Kammer) ist die kleinste Einheit im pflanzlichen und tierischen Bereich, die alle Erscheinungen des Lebens zeigt (Stoffwechsel, Wachstum, Vermehrung). In ihr laufen komplizierte Stoffwechselvorgänge ab. Sie kann sich durch Teilung vermehren. Alle höheren Lebewesen setzen sich aus einer Vielzahl von Zellen zusammen beim Menschen sind es viele Billionen -, und jede Körperzelle enthält das gesamte vom Vater und von der Mutter stammende Erbgut.
Wenngleich in Form und Funktion verschieden, z. B. Blutzellen, Drüsenzellen, Nervenzellen, Muskelzellen u. a., zeigen alle Zellen ein gleichgeartetes, hochgeordnetes Gefüge. Wir unterscheiden: Zelleib (Zytoplasma) und Zellkern (Nucleus). Die lebende, aktiv tätige Zellsubstanz heißt Protoplasma. Für die Lebensvorgänge in der Zelle spielen die Makromoleküle (Riesenmoleküle) der Eiweißkörper eine entscheidende Rolle, weil sich diese Vorgänge nur in Verbindung mit Eiweiß vollziehen können. Ohne Eiweiß gibt es kein irdisches Leben. Auf weitere Einzelheiten hinsichtlich der am Zellaufbau beteiligten Stoffe kann hier nicht näher eingegangen werden.
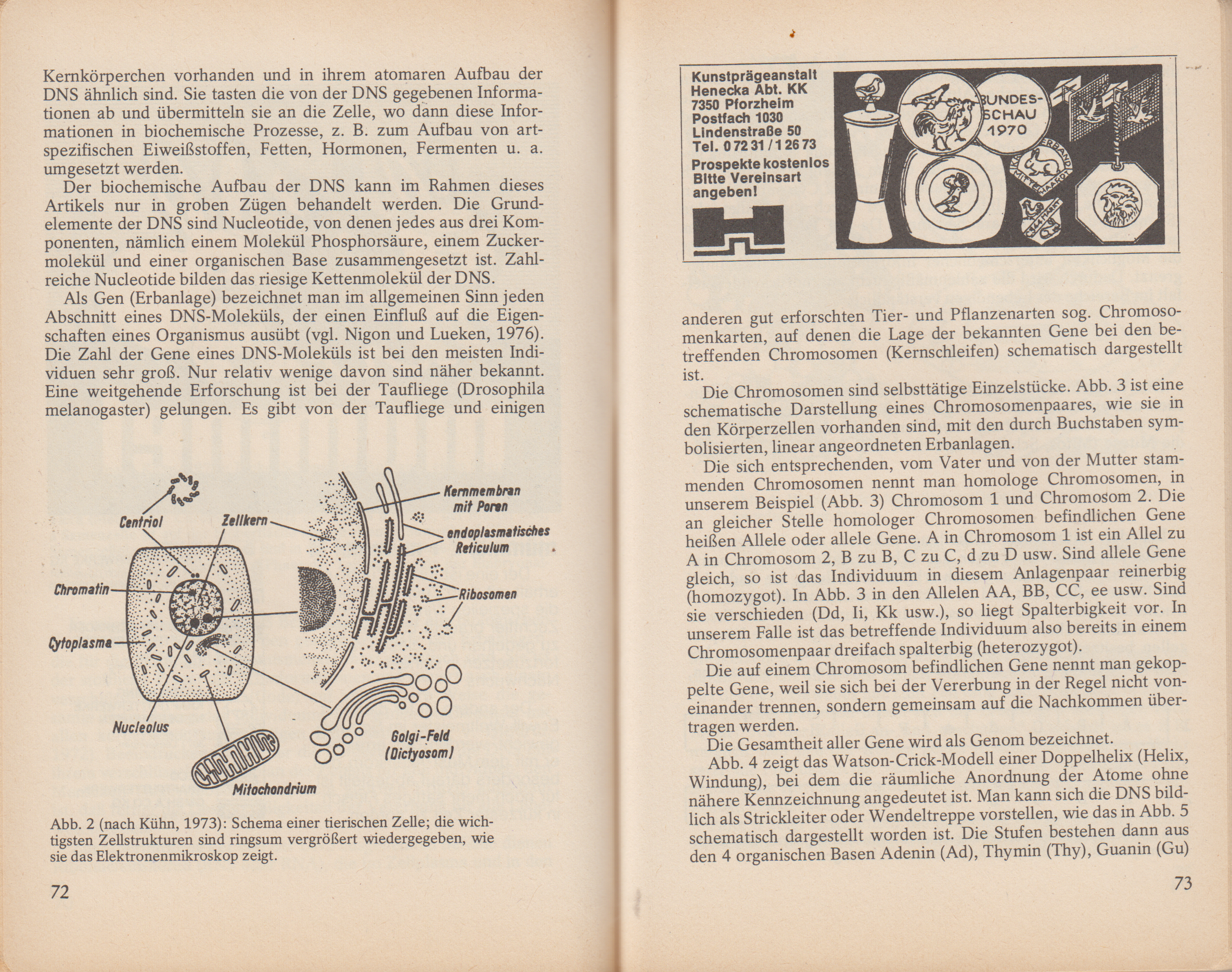
Abb. 2 zeigt eine tierische Zelle in stark vereinfachter schematischer Darstellung. Die wichtigsten Zellstrukturen sind außerhalb der Zelle nochmals stark vergrößert wiedergegeben, so wie sie unter dem Elektronenmikroskop zu erkennen sind.
Im Vordergrund unserer Betrachtung steht der Zellkern, der für alle Stoffwechselvorgänge und auch für die Vererbung eine bestimmende Rolle spielt. Er hat meist eine kugelförmige Gestalt, ist in das Zellplasma eingebettet und durch eine Doppelmembran abgegrenzt. Im Zellkern befindet sich der Kernsaft, ein oder mehrere Kernkörperchen (Nucleoli) und ein feinverteiltes Fadengerüst. Letzteres tritt besonders hervor, wenn der Zellinhalt mit Fällungsmitteln fixiert (festgemacht) und mit geeigneten Farbstoffen angefärbt wird. Da das Kerngerüst bestimmte Farbstoffe leicht aufnimmt, bezeichnet man es als Chromatin (Chroma, Farbe). Der bestimmende Bestandteil des Chromatins ist die Desoxyribonucleinsäure (DNS oder DNA), ein Wortungetüm, von dem ich im Folgenden nur die Abkürzung DNS verwenden werde.
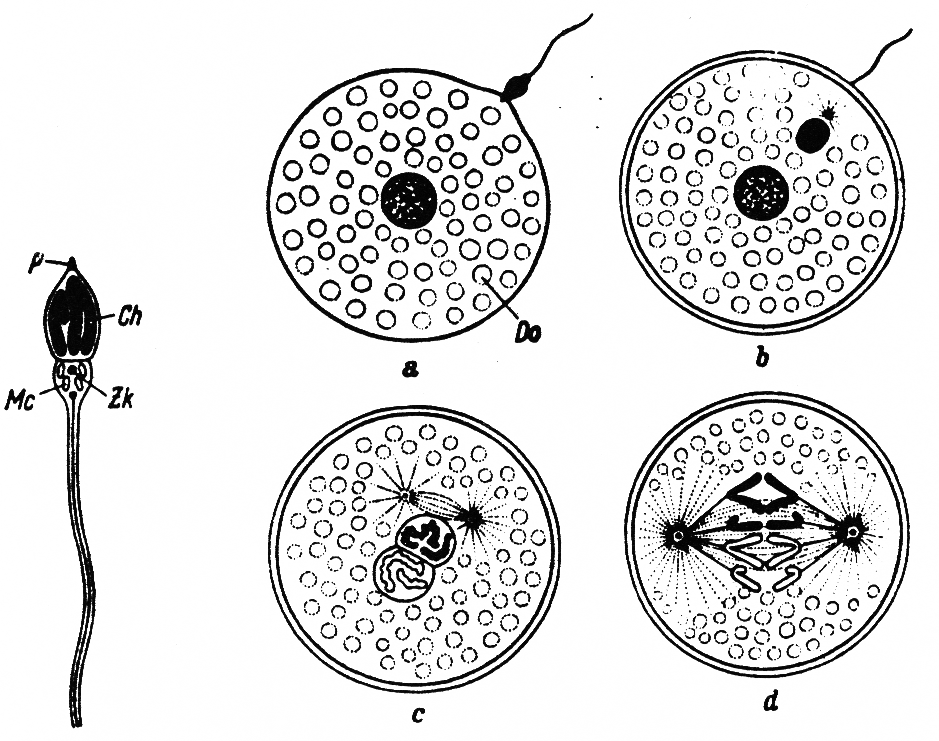
Abb. 1 (nach Kühn, 1973): Links: Schema des Aufbaues eines Spermiums; Ch Chromosomen, Mi Mitochondrien, P Perforatorium, Zk Zellkern. Rechts: schematische Darstellung des Befruchtungsverlaufs eines Eies vielzelliger Tiere (Metazoen). Chromosomensatz des Vaters schwarz, der Mutter hell; a Samenzelle ist auf der Eioberfläche aufgetroffen; b Samenkopf (Kern der Samenzelle) und Zentralkörper sind in die Zelle eingedrungen; c Verschmelzung von Ei und Samenkern; d Phase der Teilung, Beginn der Embryonalentwicklung; Do Dotter.
Es zeigte sich, dass die DNS-Befehlszentrale für alle biologischen Funktionen ist. Sie enthält den gesamten Schatz der Erbanlagen, die für die betreffende Organismenart charakteristisch sind. Sie ist der stoffliche Träger der Erbinformationen und steuert nach einem verschlüsselten Fahrplan (Code) auf wunderbare Weise die gesamte Entwicklung des Individuums und seinen Stoffwechsel mit allen körperlichen und geistigen Einzelheiten (vgl. v. Ditfurth, 1972). Beeindruckend ist ferner die Entdeckung, dass die DNS mit ihrem verschlüsselten Fahrplan des Lebens für alle Individuen, den Menschen eingeschlossen, nach dem gleichen Prinzip aufgebaut ist, dass ferner die Gesetzmäßigkeiten der Vererbung für alle Lebewesen gelten. Zur Übermittlung der von der DNS ausgehenden Befehle dienen Ribonukleinsäuren (RNS oder RNA), die im Zellplasma und in den Kernkörperchen vorhanden und in ihrem atomaren Aufbau der DNS ähnlich sind. Sie tasten die von der DNS gegebenen Informationen ab und übermitteln sie an die Zelle, wo dann diese Informationen in biochemische Prozesse, z. B. zum Aufbau von artspezifischen Eiweißstoffen, Fetten, Hormonen, Fermenten u. a. umgesetzt werden.
Der biochemische Aufbau der DNS kann im Rahmen dieses Artikels nur in groben Zügen behandelt werden. Die Grundelemente der DNS sind Nucleotide, von denen jedes aus drei Komponenten, nämlich einem Molekül Phosphorsäure, einem Zuckermolekül und einer organischen Base zusammengesetzt ist. Zahlreiche Nucleotide bilden das riesige Kettenmolekül der DNS.
Als Gen (Erbanlage) bezeichnet man im allgemeinen Sinn jeden Abschnitt eines DNS-Moleküls, der einen Einfluss auf die Eigenschaften eines Organismus ausübt (vgl. Nigon und Lueken, 1976). Die Zahl der Gene eines DNS-Moleküls ist bei den meisten Individuen sehr groß. Nur relativ wenige davon sind näher bekannt. Eine weitgehende Erforschung ist bei der Taufliege (Drosophila melanogaster) gelungen. Es gibt von der Taufliege und einigen anderen gut erforschten Tier- und Pflanzenarten sog. Chromosomenkarten, auf denen die Lage der bekannten Gene bei den betreffenden Chromosomen (Kernschleifen) schematisch dargestellt ist.
Die Chromosomen sind selbsttätige Einzelstücke. Abb. 3 ist eine schematische Darstellung eines Chromosomenpaares, wie sie in den Körperzellen vorhanden sind, mit den durch Buchstaben symbolisierten, linear angeordneten Erbanlagen.
Die sich entsprechenden, vom Vater und von der Mutter stammenden Chromosomen nennt man homologe Chromosomen, in unserem Beispiel (Abb. 3) Chromosom 1 und Chromosom 2. Die an gleicher Stelle homologer Chromosomen befindlichen Gene heißen Allele oder allele Gene. A in Chromosom 1 ist ein Allel zu A in Chromosom 2, B zu B, C zu C, d zu D usw. Sind allele Gene gleich, so ist das Individuum in diesem Anlagenpaar reinerbig (homozygot). In Abb. 3 in den Allelen AA, BB, CC, ee usw. Sind sie verschieden (Dd, Ii, Kk usw.), so liegt Spalterbigkeit vor. In unserem Falle ist das betreffende Individuum also bereits in einem Chromosomenpaar dreifach spalterbig (heterozygot).
Die auf einem Chromosom befindlichen Gene nennt man gekoppelte Gene, weil sie sich bei der Vererbung in der Regel nicht voneinander trennen, sondern gemeinsam auf die Nachkommen übertragen werden.
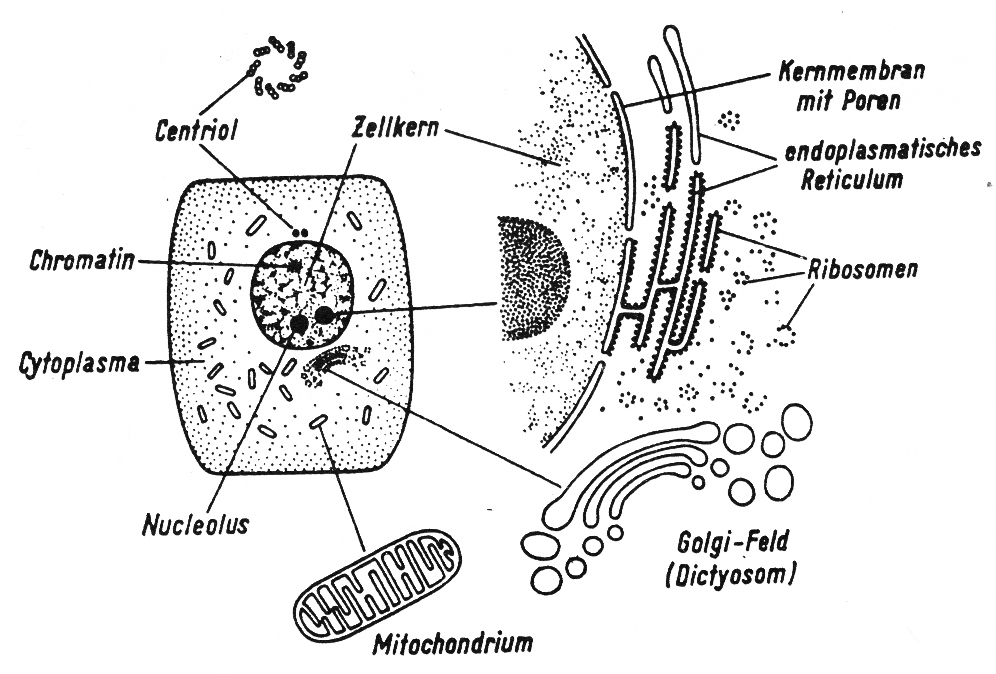
Abb. 2 (nach Kühn, 1973): Schema einer tierischen Zelle; die wichtigsten Zellstrukturen sind ringsum vergrößert wiedergegeben, wie sie das Elektronenmikroskop zeigt.
Die Gesamtheit aller Gene wird als Genom bezeichnet.
Abb. 4 zeigt das Watson-Crick-Modell einer Doppelhelix (Helix, Windung), bei dem die räumliche Anordnung der Atome ohne nähere Kennzeichnung angedeutet ist. Man kann sich die DNS bildlich als Strickleiter oder Wendeltreppe vorstellen, wie das in Abb. 5 schematisch dargestellt worden ist. Die Stufen bestehen dann aus den 4 organischen Basen Adenin (Ad), Thymin (Thy), Guanin (Gu) und Cytosin (Cyt). Die Anfangsbuchstaben (A, T, G und C) werden auch als das genetische Alphabet bezeichnet. Die Seitenteile (Holme) bestehen aus aneinandergereihten Phosphorsäure- und Zuckermolekülen (Z, Zucker; schwarze Punkte, Phosphorsäure). Dieses riesige Kettenmolekül ist in Längsrichtung spiralförmig gedreht. Jede Sprosse besteht aus einem Basenpaar, wobei sich immer nur Adenin mit Thymin und Guanin mit Cytosin verbinden können. Die Art der Information wird durch die Reihenfolge (Sequenz) der Basenpaare bestimmt. Da ein DNS-Molekül aus ungezählten Basenpaaren mit beliebiger Reihenfolge besteht, ist die Möglichkeit unterschiedlicher Informationen praktisch unbegrenzt. Dadurch wird die zahlenmäßig nicht auszudrückende Vielfalt im Bereiche des Lebendigen verständlich.
Zellteilung
Die Vermehrung der Zellen erfolgt durch Teilung. Bei Kaninchen und anderen höheren Organismen mit geschlechtlicher Fortpflanzung unterscheidet man zwei Formen der Zellteilung, nämlich die Mitose (Mitos, Schlinge) und die Meiose (Meiosis, Verminderung).
Die Mitose
Bei der mitotischen Zellteilung erhalten die Tochterzellen exakte Duplikate der Träger der Erbinformationen. Man nennt diese Form der Zellteilung auch erbgleiche Teilung. Sie findet bei den Körperzellen statt und bewirkt u. a. die Entwicklung der befruchteten Eizelle zum erwachsenen Individuum sowie den Ersatz verbrauchter Zellen bzw. Heilung bei Verletzungen. Alle Körperzellen besitzen deshalb das gleiche Erbgut. Wenn trotzdem bestimmte Zellen, z. B. Blutzellen, Nervenzellen, Drüsenzellen,
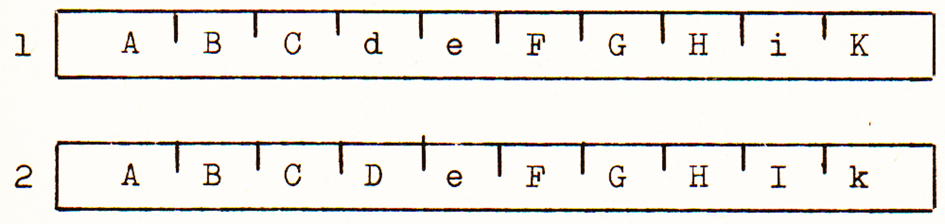
Abb. 3: Schematische Darstellung eines Chromosomenpaares. Die Buch- staben symbolisieren die linear angeordneten Erbanlagen.
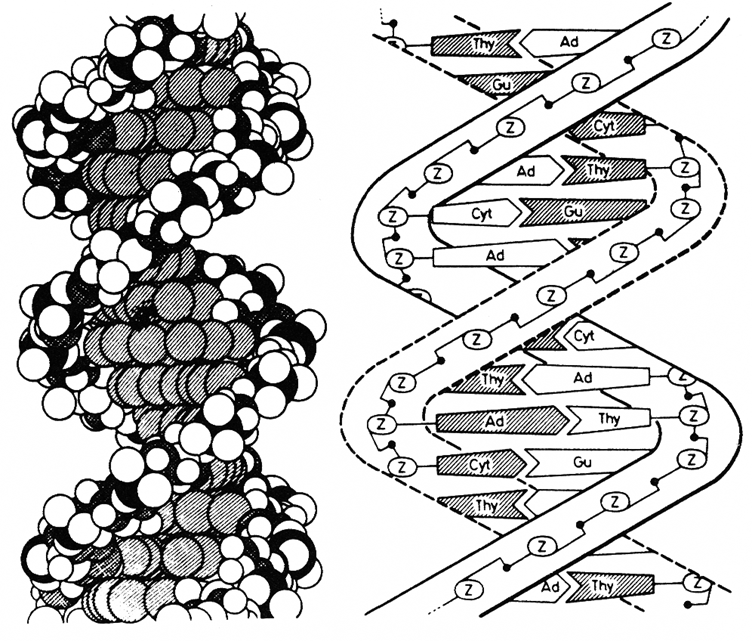
Abb. 4: Watson-Crick-Modell einer Doppelhelix mit räumlicher Anordnung der Atome.
Abb. 5 (nach Nigon und Lueken, 1976): Modell einer verdrillten Strickleiter; die Sprossen der Leiter sind durch Basenpaare gebildet; die Seitenteile (Holme) bestehen aus aneinandergereihten Phosphor- und Zuckermolekülen.
Muskelzellen u. a. unterschiedliche Aufgaben erfüllen, so liegt das daran, dass die in diesen Zellen vorhandenen Erbanlagen je nach Art des betr. Gewebes nur zum Teil tätig sind. Aber auch in Zellen mit gleicher Funktion lassen sich die Erbanlagen sozusagen nach Bedarf von der Zelle „an- und abschalten". Dadurch ist es z. B. möglich, dass sich der Körper gegen Krankheitserreger zur Wehr setzt, die Produktion der Verdauungsenzyme der Ernährungsweise anpasst, die Körpertemperatur bei unterschiedlicher Umwelt weitgehend konstant hält usw.
Abb. 6 zeigt schematisch die einzelnen Phasen der mitotischen Zellteilung. Erhält die ausgewachsene Zelle (6a) von der DNS den Befehl, sich zu teilen, so teilt sich zunächst das im Zellplasma befindliche Zentralkörperchen (Zentriol).
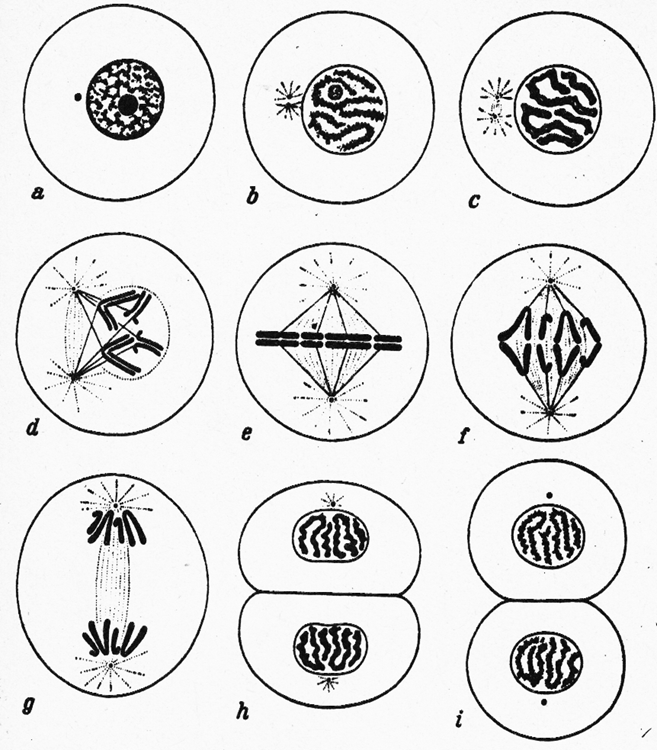
Abb. 6 (nach Kühn, 1973): Schematische Darstellung der einzelnen Phasen einer mitotischen (erbgleichen) Zellteilung.
Die beiden Teile wandern auseinander und bilden eine aus Plasmafäden bestehende Spindel (Abb. 6b, c, d und e). Gleichzeitig verdichtet sich das Kerngerüst zu den längsgespaltenen Chromosomen, die sich nach Auflösung der Kernmembran im Äquator der Spindel anordnen. Die Tochterchromosomen werden durch die Plasmafäden auseinandergezogen und bilden sich wieder zu der ursprünglichen Form des Kerngerüstes zurück. Es entsteht eine neue Kernmembran für beide Tochterkerne, der Zelleib schnürt sich durch, und beide Tochterzellen wachsen zur Größe der ursprünglichen Mutterzelle heran. Damit ist der Teilungsvorgang abgeschlossen.
Abb. 7: Chromosomen verschiedener Tierarten, wie sie sich bei Schnitten unter dem Mikroskop zeigen.
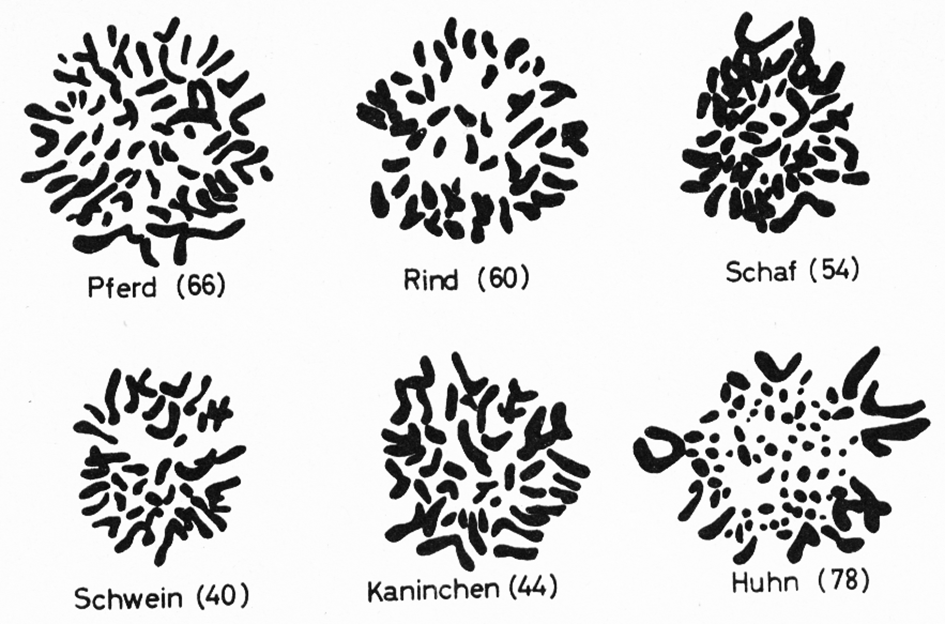
Die Zahl der Chromosomen ist bei Individuen derselben Art normalerweise gleich; sie kann jedoch von Art zu Art recht unterschiedlich sein. So hat z. B. (nach verschiedenen Autoren)
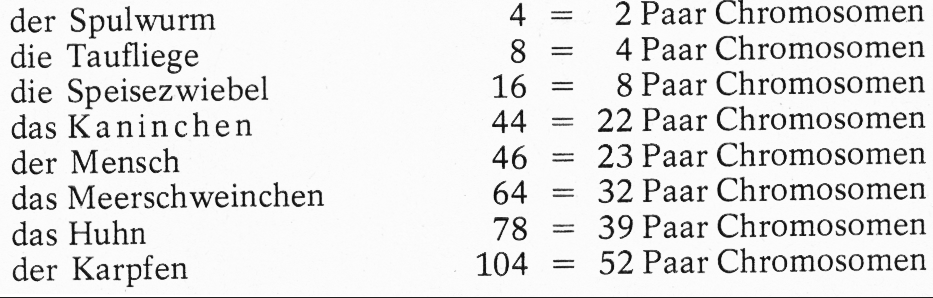
Wie die Zahlen andeuten, sind die Chromosomen in den Körperzellen in der Regel paarweise, in diploider Form, vorhanden. Jede Körperzelle besitzt zwei Chromosomensätze; der eine stammt vom Vater, der andere von der Mutter. Abb. 7 zeigt die Chromosomen verschiedener Tierarten, wie sie sich bei Schnitten unter dem Mikroskop zeigen. In Abb. 8 sind die Chromosomen des männlichen, in Abb. 9 des weiblichen Kaninchens paarweise angeordnet. Chromosomen, die beim Männchen mit XY und beim Weibchen mit XX bezeichnet werden, nennt man Geschlechtschromosomen, weil sie das Geschlecht bestimmen. Bei den meisten Tier- und Pflanzenarten findet man das ungleiche Paar (XY) beim männlichen Geschlecht, das gleiche Paar (XX) beim weiblichen. Nur bei Vögeln, Schmetterlingen und wenigen anderen Tier- und Pflanzen arten ist es umgekehrt. Die übrigen Chromosomen nennt Abb. 9:Paarweise Anordnung der Chromosomen einer Häsin; sie besitzt 2 X-Chromosomen (s. unter C).Abb. 9:Paarweise Anordnung der Chromosomen einer Häsin; sie besitzt 2 X-Chromosomen (s. unter C).Abb. 8:Paarweise Anordnung der Chromosomen eines Rammlers nach Aufnahmen durch ein Elektronenmikroskop; das X-Chromosom ist unter C, das Y-Chromosom unten rechts angeordnet.Abb. 8:Paarweise Anordnung der Chromosomen eines Rammlers nach Aufnahmen durch ein Elektronenmikroskop; das X-Chromosom ist unter C, das Y-Chromosom unten rechts angeordnet.man Autosomen.
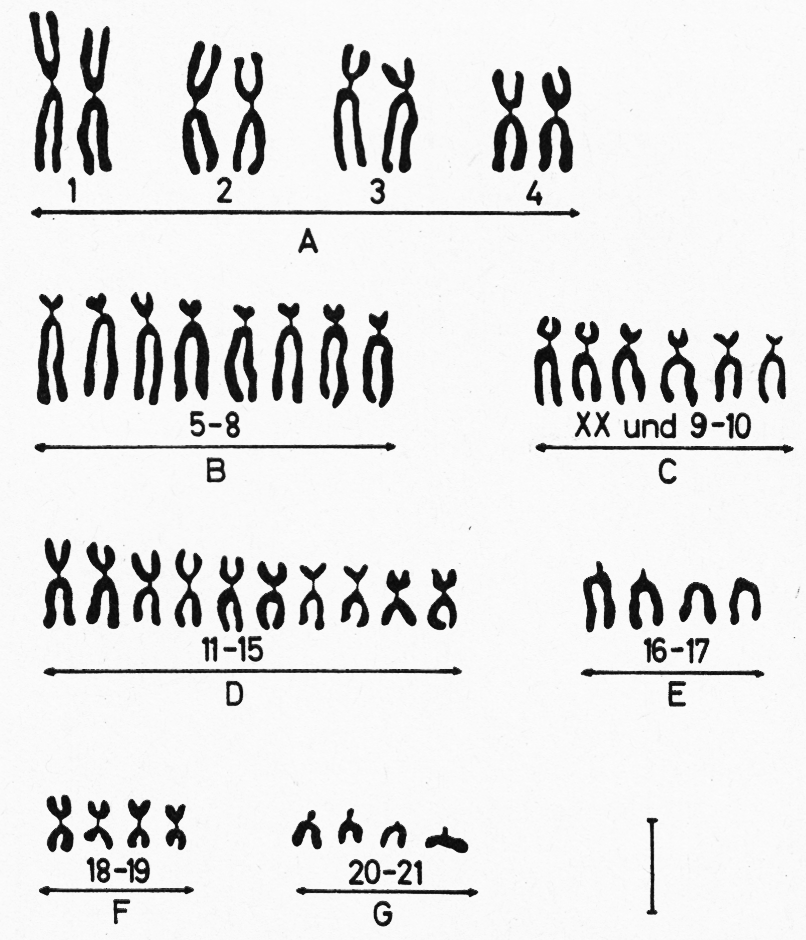

Abb. 9:
Paarweise Anordnung der Chromosomen einer Häsin; sie besitzt 2 X-Chromosomen (s. unter C).
Abb. 9:
Paarweise Anordnung der Chromosomen einer Häsin; sie besitzt 2 X-Chromosomen (s. unter C).
Abb. 8:
Paarweise Anordnung der Chromosomen eines Rammlers nach Aufnahmen durch ein Elektronenmikroskop; das X-Chromosom ist unter C, das Y-Chromosom unten rechts angeordnet.
Abb. 8:
Paarweise Anordnung der Chromosomen eines Rammlers nach Aufnahmen durch ein Elektronenmikroskop; das X-Chromosom ist unter C, das Y-Chromosom unten rechts angeordnet.
Meiose, Reifungsteilung
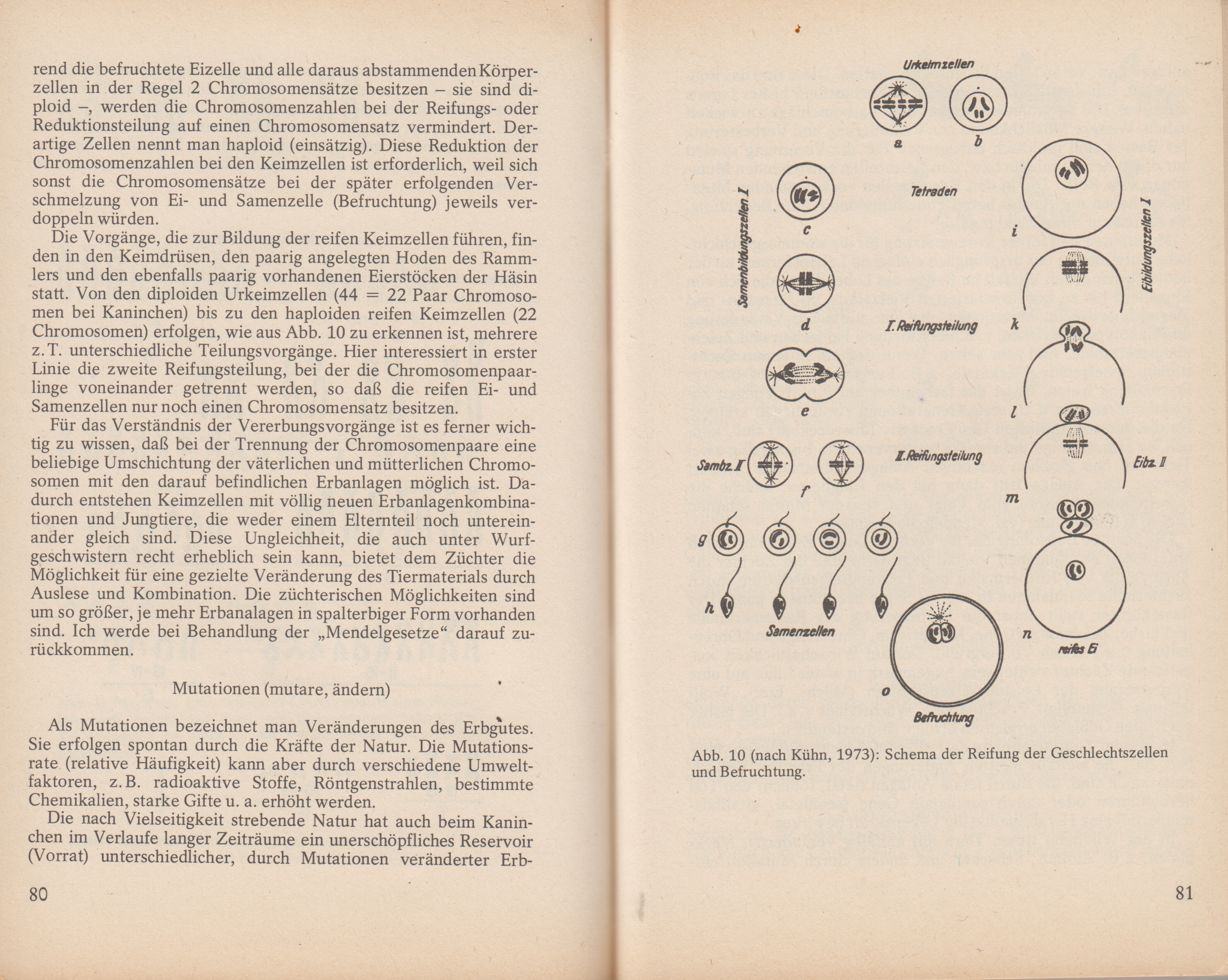
Zweck der Reifungsteilung ist die Entwicklung der Keimzellen, der männlichen Samenzellen und der weiblichen Eizellen. Während die befruchtete Eizelle und alle daraus abstammenden Körperzellen in der Regel 2 Chromosomensätze besitzen sie sind diploid, werden die Chromosomenzahlen bei der Reifungs- oder Reduktionsteilung auf einen Chromosomensatz vermindert. Derartige Zellen nennt man haploid (einsätzig). Diese Reduktion der Chromosomenzahlen bei den Keimzellen ist erforderlich, weil sich sonst die Chromosomensätze bei der später erfolgenden Verschmelzung von Ei- und Samenzelle (Befruchtung) jeweils verdoppeln würden.
Die Vorgänge, die zur Bildung der reifen Keimzellen führen, finden in den Keimdrüsen, den paarig angelegten Hoden des Rammlers und den ebenfalls paarig vorhandenen Eierstöcken der Häsin statt. Von den diploiden Urkeimzellen (44 22 Paar Chromosomen Abb. 10 (nach Kühn, 1973): Schema der Reifung der Geschlechtszellen und BefruchtungAbb. 10 (nach Kühn, 1973): Schema der Reifung der Geschlechtszellen und Befruchtungbei Kaninchen) bis zu den haploiden reifen Keimzellen (22 Chromosomen) erfolgen, wie aus Abb. 10 zu erkennen ist, mehrere z. T. unterschiedliche Teilungsvorgänge. Hier interessiert in erster Linie die zweite Reifungsteilung, bei der die Chromosomenpaarlinge voneinander getrennt werden, so dass die reifen Ei- und Samenzellen nur noch einen Chromosomensatz besitzen.

Abb. 10 (nach Kühn, 1973): Schema der Reifung der Geschlechtszellen und Befruchtung
Abb. 10 (nach Kühn, 1973): Schema der Reifung der Geschlechtszellen und Befruchtung
Für das Verständnis der Vererbungsvorgänge ist es ferner wichtig zu wissen, dass bei der Trennung der Chromosomenpaare eine beliebige Umschichtung der väterlichen und mütterlichen Chromosomen mit den darauf befindlichen Erbanlagen möglich ist. Dadurch entstehen Keimzellen mit völlig neuen Erbanlagenkombinationen und Jungtiere, die weder einem Elternteil noch untereinander gleich sind. Diese Ungleichheit, die auch unter Wurfgeschwistern recht erheblich sein kann, bietet dem Züchter die Möglichkeit für eine gezielte Veränderung des Tiermaterials durch Auslese und Kombination. Die züchterischen Möglichkeiten sind umso größer, je mehr Erbanalagen in spalterbiger Form vorhanden sind. Ich werde bei Behandlung der „Mendelgesetze" darauf zurückkommen.
Mutationen (mutare, ändern)
Als Mutationen bezeichnet man Veränderungen des Erbgutes. Sie erfolgen spontan durch die Kräfte der Natur. Die Mutations- rate (relative Häufigkeit) kann aber durch verschiedene Umweltfaktoren, z. B. radioaktive Stoffe, Röntgenstrahlen, bestimmte Chemikalien, starke Gifte u. a. erhöht werden. Die nach Vielseitigkeit strebende Natur hat auch beim Kaninchen im Verlaufe langer Zeiträume ein unerschöpfliches Reservoir (Vorrat) unterschiedlicher, durch Mutationen veränderter Erbanlagen bzw. veränderten Erbgutes geschaffen. Diese sind das Roh- material, mit dem Züchterkönnen und Züchterfleiß bisher bereits weit über 100 verschiedene Rassen und Farbenschläge entwickelt haben. Weitere Möglichkeiten zur Veränderung und Verbesserung der Rassen sind praktisch unbegrenzt. Für die Vererbung spielen nur die in der Keimbahn bzw. den Keimzellen entstehenden Mutationen eine Rolle. Die in den Körperzellen vorkommenden Mutationen haben nur für das betreffende Individuum eine Bedeutung. Sie führen zur Mosaikbildung.
Abb. 11 (in Anlehnung an verschiedene Autoren): Schematische Darstellung der Variationsbreite des Scheckungsmusters bei der Englischen Scheckung, bei der offenbar zahlreiche modifizierende Gene eine Rolle spielenAbb. 11 (in Anlehnung an verschiedene Autoren): Schematische Darstellung der Variationsbreite des Scheckungsmusters bei der Englischen Scheckung, bei der offenbar zahlreiche modifizierende Gene eine Rolle spielen Mutationen sind ferner Voraussetzung für die stammesgeschichtliche Entwicklung der ursprünglich einfachen Lebensformen zu der heute vorhandenen Vielfalt im Reiche des Lebendigen, die sich von Einzellern bis zu hochorganisierten Vielzellern im Pflanzen- und Tierreich erstreckt. Mutationen, die eine auffällige Veränderung im Phänotypus bewirken, z. B. verschiedene Fellfarben und Zeichnungsmerkmale, sind sehr selten. Wenn deshalb in einem Zuchtstamm rassefremde Merkmale, z. B. Langhaar, Albinos u.a. in Erscheinung treten, so ist das fast immer auf Einkreuzungen zu- rückzuführen, die u. U. viele Generationen zurückliegen können. Da die meisten mutierten Gene rezessiv (überdeckbar) sind, können sie sich so lange unbemerkt weitervererben, bis zufällig zwei Tiere mit den gleichen rezessiven Anlagen miteinander gepaart werden. Die Anlage tritt dann bei den Jungtieren, welche die rassefremden Anlagen vom Vater und von der Mutter erhalten haben, in Erscheinung (vgl. Mendelsche Gesetze). Die positive Bedeutung der im Verlaufe langer Zeiträume durch Mutationen entstandenen Vielfalt liegt darin, dass der Züchter die Ausprägung von Körpermalen und Leistungseigenschaften durch züchterische Maßnahmen in seinem Sinne beeinflussen kann. Der Rassezüchter stellt dabei die Verbesserung der Rassemerkmale (Fellfarbe, Zeichnung, Größe, Körperform, Ohrenlänge und Ohrenhaltung u. a.) in den Vordergrund. Der auf Wirtschaftlichkeit aus- gerichtete Züchter richtet sein Augenmerk in erster Linie auf eine Verbesserung der Leistungseigenschaften (Milch-, Eier-, Wollleistung, Gesundheit, Fruchtbarkeit, Wüchsigkeit u. a.). Die bisher auf beiden Gebieten erreichten Erfolge sind erstaunlich. In diesem Zusammenhang darf allerdings nicht unerwähnt bleiben, dass die weitaus meisten Änderungen des Erbgutes Verlustmutationen sind, die durch letale Anlagen (letal, tödlich) den Tod herbeiführen oder durch semiletale Gene (semiletal, vitalitätsmindernd) eine Herabsetzung der Lebenskraft bewirken. In der Wildbahn treten Tiere mit auffällig veränderten Merkmalen, z. B. Albinos, Schecken und andere durch Mutation entstandene Typen nur selten in Erscheinung, obwohl sich auch bei Wildtieren viele mutierte Gene angesammelt haben. Das liegt einmal daran, dass es sich in den weitaus meisten Fällen um rezessive Gene handelt, die nur in reinerbiger Form in Erscheinung treten, und andererseits die reinerbig behafteten Tiere den phänotypisch nicht veränderten Artgenossen im Kampf ums Dasein unterlegen sind.
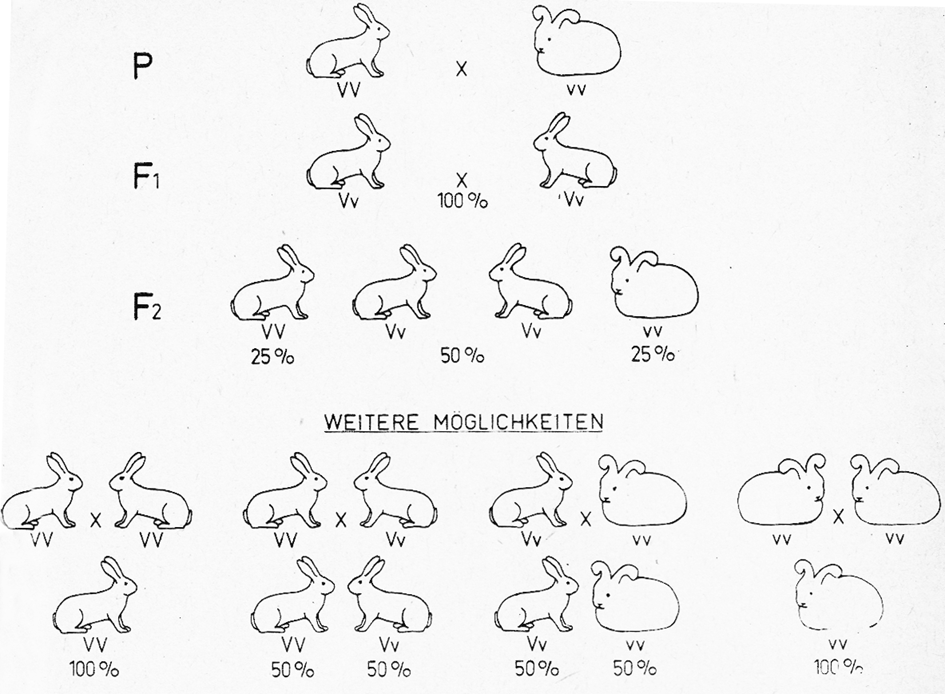
Abb. 11 (in Anlehnung an verschiedene Autoren): Schematische Darstellung der Variationsbreite des Scheckungsmusters bei der Englischen Scheckung, bei der offenbar zahlreiche modifizierende Gene eine Rolle spielen
Abb. 11 (in Anlehnung an verschiedene Autoren): Schematische Darstellung der Variationsbreite des Scheckungsmusters bei der Englischen Scheckung, bei der offenbar zahlreiche modifizierende Gene eine Rolle spielen
Formen der Mutationen
Genmutationen entstehen durch biochemische Veränderungen der DNS im Bereiche eines Gens bzw. Triplets. Nur wenige bewirken auffallende Veränderungen im Phänotypus, z. B. unterschiedliche Fellfarben und Zeichnungsmerkmale. Die meisten Mutationen sind äußerlich nicht zu erkennen und als solche nur schwer zu identifizieren.
Chromosomenmutationen
verändern die lineare Anordnung der Chromosomenabschnitte. Das kann durch Chromosomenbrüche geschehen, bei denen Bruchstücke verlorengehen oder wechselseitig ausgetauscht werden. Ein solcher Faktorenaustausch kann sich auch positiv auswirken, wenn dadurch unerwünschte Gene von im selben Chromosom befindlichen erwünschten getrennt werden. Möglicherweise ist das u. a. beim Rexkaninchen geschehen, das ursprünglich neben dem gewünschten Kurzhaar negative Erscheinungen (Konstitutions- schwächen, erhöhte Sterblichkeit der Jungen, schlechte Futterverwertung, Anfälligkeit gegen Erkrankungen der Luftwege, Augenkrankheiten u. a.) zeigte. Heute sind diese negativen Erscheinungen fast völlig verschwunden. Natürlich ist ein Faktorenaustausch nicht die einzige Möglichkeit einer Vitalitätsverbesserung.
Genommutationen
Dabei bleiben die einzelnen Chromosomen und die darauf befindlichen Gene unverändert. Von den verschiedenen Möglichkeiten der Genommutationen sei hier in erster Linie die Euploidie erwähnt, bei der der Chromosomensbestand einer Zelle um ganze Chromosomensätze vermehrt oder vermindert wird. Das Auftreten mehrerer Chromosomensätze bezeichnet man als Polyploidie (polys, viel). Die meisten Genommutationen kommen durch Hemmung des normalen Kernteilungsprozesses zustande. Sie können auch experimentell ausgelöst werden, wobei das Spindelgift der Herbstzeitlose, das Colchicin, einer der auslösenden Faktoren ist. Vor mehreren Jahren wurde berichtet, dass es in Schweden gelungen sei, ein triploides Kaninchen (3 Chromosomensätze) zu erzeugen. Da man sich später darüber ausschwieg, ist anzunehmen, dass eine Weiterzüchtung nicht gelungen ist. Genommutationen, die in der Pflanzenwelt eine große Rolle spielen, haben bei Tieren – soweit mir bekannt ist – bisher meist nur negative Auswirkungen gezeigt.
Modifikationen
sind umweltbedingte, nicht erbliche Veränderungen des Phänotypus. Die phänotypische Entwicklung eines Organismus wird durch das Zusammenwirken von Erbanlagen und Umwelt bestimmt. Die Erbanlagen legen dabei die Grenzen fest (Reaktionsnorm), innerhalb derer die Umweltfaktoren wirksam werden können. Zu den wichtigsten Umweltfaktoren gehören Ernährung, Haltung, Krankheitserreger, Temperatur, Luftfeuchtigkeit, Stressfaktoren u. a. Jedem erfahrenen Züchter ist bekannt, dass man z. B. durch Maßnahmen der Fütterung Wachstum, Gesundheit, Fruchtbarkeit, Milch- und Wolleistungen u. a. erheblich beeinflussen kann. Ausgewogene Fütterung fördert, Fütterungsfehler hemmen die Entwicklung der Leistungseigenschaften. Die durch Umwelteinflüsse bedingten Unterschiede, die nicht auf die Nachkommen übertragen werden, können ganz oder teilweise bei den betreffenden Tieren noch ausgeglichen werden, wenn man die Ursachen beseitigt. So können z. B. Jungtiere mit guter Veranlagung, die bei großen Würfen infolge ungenügender Milchaufnahme in der Entwicklung zurückgeblieben sind, den Rückstand später bei ausgewogener Fütterung in relativ kurzer Zeit wieder aufholen. Deshalb trifft auch die Meinung mancher Züchter, gute Ausstellungstiere könnten nur aus kleinen Würfen (3-4 Jungtiere) entwickelt werden, nicht zu. Wer seine Würfe unnötig reduziert oder nur Tiere von kleinen Würfen aufzieht, handelt sich dabei erhebliche Nachteile ein:
1. Bei Verwendung von Jungtieren aus kleinen Würfen für die Ausstellung und Nachzucht erreicht man über kurz oder lang eine verminderte Fruchtbarkeit, was u. U. sogar zum Versagen in der Zucht führen kann;
2. verringert man die Möglichkeit einer Auswahl – vielleicht hat man die besten Tiere ausgemerzt- und erhöht das Risiko, bei eventuellen Jungtierverlusten nicht genügend wertvolle Ausstellungs- und Zuchttiere zur Verfügung zu haben.
Der Umwelteinfluss auf die verschiedenen Merkmale und Eigenschaften ist recht unterschiedlich. Er wirkt sich am stärksten bei den Leistungseigenschaften, am geringsten bei den Körpermerkmalen, besonders der Fellfarbe und Zeichnung aus. So ändern sich die Zeichnungsmerkmale der Schecken (Engl. Schecken, Holländerschecken) nach der Geburt der Tiere kaum noch, so dass der Züchter eine Vorauslese der gutgezeichneten Tiere bereits kurz nach der Geburt vornehmen kann.
Der Umwelteinfluss ist aber auch bei den Leistungseigenschaften nicht unbegrenzt. Er kann sich nur innerhalb der von den Erbanlagen festgelegten Grenzen (Reaktionsnorm) auswirken. So kann man trotz Optimierung aller Umweltfaktoren aus einem Zwergkaninchen keinen Riesen machen. Auch umgekehrt lassen sich Kaninchen mit Anlagen für Riesen nicht beliebig klein halten, ohne dass schwere gesundheitliche Schäden auftreten. Auch die Wollleistungen der Angorakaninchen sind, wie jeder erfahrene Angorazüchter weiß, durch die jeweils vorhandenen Erbanlagenkombinationen begrenzt. Das erhärten auch die großen Leistungsstreuungen, die bei den unter möglichst gleichen Bedingungen auf den ALP-Stationen gehaltenen Prüflingen sichtbar werden. Auch eigene langjährige Erfahrungen in der Angorazucht lassen daran keinen Zweifel.
Gensymbole
Mutationen mit im Erscheinungsbild sichtbaren Wirkungen bieten die Möglichkeit, das Erbgut zu zergliedern und auf das Vorhandensein entsprechender Erbanlagen zu schließen. Wenn z. B. ein Tier in Erscheinung tritt, dass sich vom Wildkaninchen durch eine blaue, braune, gelbe oder schwarze Fellfarbe oder durch bestimmte Zeichnungsmerkmale unterscheidet, so ist dem Fachmann klar, dass sich beim wildfarbigen Kaninchen entsprechende allele Gene befinden und dass die Änderung der Fellfarbe auf eine Mutation dieser Gene zurückzuführen ist. Wir wollen uns hier auf Gene mit einfachen Erbgängen beschränken.
Um damit Erbanalysen vornehmen und die statistisch (im Durch- schnitt großer Zahlen) zu erwartenden Aufspaltungserscheinungen zahlenmäßig besser erfassen zu können, hat man die bekannten Gene durch Buchstaben als Symbole gekennzeichnet. Ich werde dabei die von Prof. Dr. Hans Nachtsheim benannten Symbole verwenden, möchte dazu aber bemerken, dass in der englischsprachigen Literatur für die gleichen Gene andere Symbole benutzt werden. Eine ausführliche Erläuterung hinsichtlich der Wirkungsweise der verschiedenen Gene mit konkreten Beispielen für die Durchführung auch schwieriger Erbanalysen würde den Rahmen dieses Artikels sprengen. Ich möchte mich hier auf einige Hinweise beschränken.
Die Genformel für die Fellfarbe aller wildgrauen Kaninchenrassen lautet: ABCDG. Streng genommen müsste es heißen: AABBCCDDGG, weil ja alle Gene paarweise vorhanden sind. Aus Gründen einer besseren Übersicht werden Gene, die in reinerbiger Form vorhanden sind, nur einfach aufgeführt. Bei Spalterbigkeit müssen die betreffenden Genpaare natürlich vollständig angegeben werden, z. B. Aa, Bb usw. Für Erbanalysen benutzt man zur Vereinfachung nur diejenigen Gensymbole, die bei dem betreffenden Erbvorgang eine Rolle spielen. Ich werde bei später aufgeführten Beispielen noch darauf zurückkommen.
Sollen bei der Erbanalyse außer den für die Fellfarbe verantwortlichen Genen noch andere, z. B. für die verschiedenen Zeichnungsmerkmale, für die Silberung, Haarlänge und Haarstruktur, Erbkrankheiten u. a. zuständigen Erbanlagen untersucht werden, so muss man die entsprechenden Gensymbole natürlich einbeziehen.
Von den in der obigen Erbformel aufgeführten Gensymbolen ist A der Grundfaktor für eine Pigmentierung. Eine ähnliche Wirkung hat der Faktor X, der aber meist nur dann aufgeführt wird, wenn er bei der Erbanalyse eine Rolle spielt.
Findet eine Mutation von A zu a statt, so ist bei den reinerbigen aa-Tieren eine Pigmentbildung nicht möglich. Es entstehen Albi- nos, Tiere mit weißem Fell und roten Augen. Bei einer Mutation von X zu x haben die xx-Tiere ebenfalls ein weißes Haarkleid. Es erfolgt aber noch eine Pigmentierung der Iris (Regenbogenhaut). Die Augenfarbe erscheint blau. B, C und D sind die eigentlichen Farbfaktoren. G ist der Farbverteilungsfaktor, der die zonenweise Aufteilung der Pigmente bewirkt, wie das am deutlichsten bei den wildgrauen und chinchillafarbigen Rassen in Erscheinung tritt. Eine Mutation von G zu g bewirkt schwarze Fellfarbe, von D zu d blaue, von C zu c braune und von B zu b gelbe Fellfarbe. Die Zeichnungsrassen, die Kurzhaarrassen sowie das langhaarige Angorakaninchen sind ebenfalls durch Mutationen entstanden.
Die dominanten (überdeckenden) Erbanlagen werden im all- gemeinen mit großen Buchstaben, die rezessiven (überdeckbaren) Allele mit kleinen Buchstaben symbolisiert. So ist z. B. A dominant über a, B über b usw. Hiervon gibt es allerdings Ausnahmen. Das trifft für polyallele, das sind mehrfach mutierte Gene, zu.
Beispiele (nach Nachtsheim, 1977):
Die Dominanz erfolgt bei den aufgeführten Beispielen von oben nach unten. In der Albinoserie ist A über alle darunter stehenden Allele dominant. Dasselbe gilt für achi usw. Hier kommt es also vor, dass Gene mit kleinen Buchstaben über andere mit ebenfalls kleinen Buchstaben dominant sind. In der B-Serie gilt zwischen bj (Japaner) und b (gelb), ferner in der G-Serie für lohfarbig (go) und nichtwildfarbig (g) dasselbe. Im oberen Teil der B-Serie gibt es eine Dominanz von Bee über Be und B. Hier sind also auch Gene mit großen Buchstaben dominant über andere mit ebenfalls großen Buchstaben. Umgekehrt ausgedrückt können auch Gene mit großen Buchstaben rezessiv sein. Es gibt auch Fälle, in denen bestimmte Gene über nicht allele Gene dominieren. So werden z. B. alle Farb- und Zeichnungsgene unwirksam, wenn der nicht allele Albinofaktor a in reinerbiger Form (aa) vorhanden ist. Ferner wird durch achi das durch einen nicht allelen Faktor bewirkte gelbe Pigment unwirksam. Man nennt in solchen Fällen die unterdrückenden Gene epistatisch, die unterdrückten hypostatisch. In fast allen Fällen wird die Wirksamkeit eines rezessiven Gens durch das dominante Allel nicht völlig wirkungslos. Man spricht dann von unvollständiger Dominanz.
Beispiele:
Der spalterbige Typenmarder (aman oder ama) ist stärker aufgehellt und zeigt die Marderzeichnung klarer als der reinerbige Dunkelmarder (amam). Besonders deutlich ist das auch bei Englischen Schecken zu erkennen. Die Zeichnungsmerkmale (pigmentierte Stellen) treten bei den spalterbigen Typenschecken (kk) wesentlich stärker in Erscheinung als bei den reinerbigen Weißlingen (KK). Ist die Wirkung von allelen Genen gleich, so spricht man von einer intermediären (dazwischenliegenden) Vererbung. Ein klassisches Beispiel ist die Wunderblume (Mirabilis jalapa; Erbgang s. Mendelgesetze).
Es sei noch erwähnt, dass spalterbige Individuen auch Hybriden oder Bastarde genannt werden.
Monohybride Individuen sind nur in einem Anlagenpaar, dihybride zweifach, trihybride dreifach, polyhybride vielfach spalterbig.
Polygene Bestimmung der Merkmale
Alle Merkmale werden durch das Zusammenwirken vieler Erbanlagen bestimmt. Diese vielseitige Bestimmung eines Merkmals kann sich in mannigfaltiger Weise vollziehen.
1. Das wildfarbige Haarkleid des Kaninchens kommt durch das Zusammenwirken vieler Erbfaktoren zustande. Einige davon (ABCDG) wurden bereits erwähnt. Auch auf die Veränderung des Erscheinungsbildes durch Mutation eines oder mehrerer dieser Gene wurde bereits hingewiesen. Der Erbgang solcher Merkmale ist relativ leicht zu analysieren (s. Mendelgesetze).
2. In vielen Fällen, z. B. beim Zustandekommen der rostroten Farbe der Roten Neuseeländer, der Lohe bei Schwarzloh, besonders aber bei den Leistungseigenschaften (Wolleistung, Milchleistung u.a.) verstärken Gene mit gleichsinniger Wirkung den Ausbildungsgrad, d. h. je mehr derartige Gene vorhanden sind, desto stärker ist die Wirkung. Man spricht hier von einer additiven Genwirkung, weil sich die Wirkung solcher Gene addiert und damit insgesamt verstärkt. Da es sich um viele nach Zahl und Wirkung nicht näher bekannte Gene handelt, kann der Erbgang nicht durch die Mendelgesetze, sondern nur durch komplizierte mathematisch-statistische Methoden analysiert werden.
3. Häufig gibt es neben „Hauptgenen", deren Wirksamkeit sich auf ein Merkmal bezieht, z. B. Fellfarbe und Zeichnung, sog. modifizierende „Nebengene“ (modifizierend, verändernd), die den Wirkungsgrad eines „Hauptgens“ verstärken oder hemmen.
Mutiert z. B. das mit D symbolisierte Gen zu d, so zeigen die reinerbigen dd-Tiere eine blaue Fellfarbe. Die Intensität der Fell- farbe, die von Hellblau mit allen Übergängen zu Dunkelblau variieren kann, wird vermutlich durch eine große Zahl modifizierender Gene bewirkt. Das gleiche gilt sinngemäß auch für alle anderen Fellfarben.
Ein weiteres Beispiel liefert das Scheckungsmuster bei Punktscheckung (Englische Schecken, Deutsche Riesenschecken, Rheinische Schecken, Dalmatinerschecken, Deutsche Widderschecken u. a.). Die Scheckung selbst wird durch die unvollständig dominante Erbanlage K bewirkt, die in reinerbiger Form (KK) nur schwach gezeichnete Weißlinge, in spalterbiger Form (Kk) die im Standard anerkannten Typenschecken bewirkt. Der Aufhellungsgrad ist wegen der unvollständigen Dominanz von K bei den reinerbigen Weißlingen stärker als bei den spalterbigen Typenschecken. Abgesehen davon zeigt das Zeichnungsmuster aber auch bei spalterbigen Typenschecken eine erhebliche Streubreite. In Abb. 11 sind einige der dabei möglichen Muster schematisch dar- gestellt. Züchter der obengenannten Rassen wissen, dass die Natur noch wesentlich mehr Varianten hervorbringt als im Schema angegeben. Diese Vielzahl der möglichen Scheckungsmuster wird durch eine offenbar große Zahl modifizierender Gene ermöglicht.
Deshalb ist es auch so schwierig, einigermaßen konstant vererbende Schecken zu züchten. Auch bei den Holländerschecken, bei denen das Zustandekommen unterschiedlicher Scheckungsmuster noch mit einigen Fragezeichen versehen werden muss, spielen neben den Holländerfaktoren (S1, S2, S3 . . .) zweifellos auch modifizierende Gene eine Rolle. Es gibt noch viele weitere Beispiele, auf deren Darstellung hier verzichtet werden muss.
Vererbung des Geschlechts
Abb. 12: Der geniale Augustinerpater Johann Gregor Mendel, dessen fundamentale Erkenntnisse auf dem Gebiete der Vererbung von seinen Zeit- genossen nicht erkannt wurden, heute aber noch die Grundlage der Vererbung bilden.Abb. 12: Der geniale Augustinerpater Johann Gregor Mendel, dessen fundamentale Erkenntnisse auf dem Gebiete der Vererbung von seinen Zeit- genossen nicht erkannt wurden, heute aber noch die Grundlage der Vererbung bilden.Wie bereits erwähnt, wird das Geschlecht bei Kaninchen und anderen getrenntgeschlechtlichen Organismen durch die Wirkung der Geschlechtschromosomen bestimmt (XY beim Rammler, XX bei der Häsin). Da bei der Reduktionsteilung 50% der Spermien ein X- und 50% ein Y-Chromosom enthalten, alle Eizellen aber ein X-Chromosom besitzen, müssten nach den Regeln der Wahrscheinlichkeit 50% der Eizellen von einem männlich bestimmen- den Y-Spermium und 50% von einem weiblich bestimmenden X-Spermium befruchtet werden. Das entspricht auch im Durchschnitt großer Zahlen im Wesentlichen den Realitäten, wobei allerdings in Abhängigkeit von den verschiedenen Arten Abweichungen nach der einen oder anderen Seite vorkommen. So beträgt nach Dietrich, Stöcker et al (1976) das Verhältnis von Weibchen zu Männchen bei der Maus 100:79, beim Pferd und Schaf 100:98, beim Hausschwein 100:102, bei der Ratte 100:107, beim Hund 100:117 und beim Menschen 100:105. Nach eigenen Erfahrungen überwiegen auch bei Nerzen und Kaninchen die männlichen Nachkommen, wobei ich allerdings keine statistisch abgesicherten Zahlenverhältnisse nennen kann. Als Ursachen für die von 1:1 ab weichenden Werte werden genannt: Schnellere Bewegung der etwas leichteren männlichen Spermien, unterschiedliche Widerstandsfähigkeit der verschiedenen Keimzellen oder der befruchteten Eizellen gegen äußere Einflüsse (Temperatur, Chemikalien, Säuregrad u. a.). So ist z. B. bekannt, dass beim Menschen ein erheblicher Teil, besonders der männlichen Keime, auf einer frühen Entwicklungsstufe abstirbt. Auch beim Kaninchen ist die nur etwa 70% betragende Trächtigkeitsrate vermutlich u. a. auf einen embryonalen Frühtod zurückzuführen. Ein Einfluss der Geschlechtschromosomen ist dabei allerdings nicht bekannt.

Abb. 12: Der geniale Augustinerpater Johann Gregor Mendel, dessen fundamentale Erkenntnisse auf dem Gebiete der Vererbung von seinen Zeit- genossen nicht erkannt wurden, heute aber noch die Grundlage der Vererbung bilden.
Abb. 12: Der geniale Augustinerpater Johann Gregor Mendel, dessen fundamentale Erkenntnisse auf dem Gebiete der Vererbung von seinen Zeit- genossen nicht erkannt wurden, heute aber noch die Grundlage der Vererbung bilden.
Geschlechtskontrollierte Vererbung
Man versteht darunter die Erscheinung, dass sich bestimmte Merkmale und Eigenschaften im männlichen und weiblichen Geschlecht in unterschiedlicher Weise äußern. Das gilt z. B. für die Geschlechtsorgane der Rammler und Häsinnen (primäre Geschlechtsmerkmale) und auch für den Geschlechtscharakter, z. B. Unterschiede im Körperbau, im Temperament u. a. (sekundäre Geschlechtsmerkmale). Diese Unterschiede beruhen auf hormonalen Wirkungen, die von den Geschlechtschromosomen gesteuert werden. Nach Gleichauf, 1972, sind daran auch autosomale Gene beteiligt. Die komplizierten Zusammenhänge mit ihren Besonderheiten, z. B. Zwitterbildung, Kryptorchismus (unterbliebener Hodenabstieg) u. a. können hier nicht näher behandelt werden.
Geschlechtsgebundene Vererbung
Die Geschlechtschromosomen sind nicht nur für die Bestimmung des Geschlechtes verantwortlich. Speziell die X-Chromosomen enthalten auch Anlagen, die die Ausbildung von Körpermerkmalen beeinflussen. Als Beispiel sei die Bluterkrankheit (Hämophilie) beim Menschen genannt, die durch ein rezessives, im X-Chromosom befindliches Gen hervorgerufen wird. Dadurch wird die Blutgerinnung gehemmt, so dass selbst kleinste Verletzungen Lebensgefahr bedeuten, wenn der Patient nicht rechtzeitig behandelt wird. Interessant ist hierbei die Tatsache, dass diese Krankheit nur bei Männern auftritt, aber über Frauen vererbt wird. Auf eine ausführliche Darstellung des Erbganges muss hier aus Platzgründen verzichtet werden. Entsprechende Beispiele beim Kaninchen sind mir nicht bekannt.
Plasmatische oder Extrachromosomale Vererbung
Dabei handelt es sich um Vererbungsvorgänge, an denen vorwiegend oder ausschließlich im Zellplasma befindliche, also nicht- chromosomale Erbträger beteiligt sind. Der Erbgang der betreffenden Merkmale folgt nicht den Mendelgesetzen. Reziproke Kreuzungen ergeben unterschiedliche Nachkommen, da das Zytoplasma (Zellplasma) im Wesentlichen über die mütterliche Seite weitergegeben wird (Dietrich, Stöcker et al, 1976). Ab 1960 gelang es, DNS in den im Zellplasma lokalisierten Mitochondrien (Abb. 2) und Plastiden (in pflanzlichen Zellen) nachzuweisen und zu zeigen, dass diese ihre Erbinformationen mehr oder weniger unabhängig von den Chromosomen abgeben.
Die Wirkung von Plasmafaktoren ist u. a. bei bestimmten Mikroorganismen (Chlamydomonas) untersucht und dabei festgestellt worden, dass diese die Resistenz (erbliche Widerstandsfähigkeit) gegenüber zahlreichen Antibiotika kontrollieren (Geissler, Libbert, Nitschmann und Petersein, 1978).
Abb. 13: Schema eines monohybriden Erbganges am Beispiel einer Kreuzung zwischen Langhaar und Normalhaar (P); die F1-Tiere sind alle normalhaarig; in der F2 zeigt sich eine Aufspaltung Normalhaar zu Langhaar von 3:1; darunter sind weitere Möglichkeiten dargestellt.

Die Mendelschen Gesetze
Johann Gregor Mendel (Abb. 12), der sich seit 1856 mit botanischen Arbeiten beschäftigte, leitete aus seinen Kreuzungsversuchen, die er mit Erbsen, Bohnen, Fuchsien und Kapuzinerkresse im Klostergarten von Brünn durchführte, die Mendelschen Gesetze für die Vererbung einfacher Merkmale ab. Als Ausgangsmaterial verwendete er durch Selbstbestäubung gezüchtete reine Rassen mit unterschiedlichen Merkmalen, die er miteinander kreuzte. Aus den Ergebnissen dieser Versuche hatte Mendel als erster erkannt, dass nicht die Merkmale, sondern die dafür verantwortlichen Erbanlagen auf die Nachkommen übertragen werden, dass ferner das Erbgut aus vielen unabhängig voneinander vererbenden Anlagen besteht und diese von der Weitergabe an die Nachkommen nach bestimmten Gesetzmäßigkeiten umgruppiert werden. Die dadurch hervorgerufene Verschiedenheit (Variation) bietet die Möglichkeit einer Auslese (Selektion) und die Voraussetzung, bis- her bei verschiedenen Individuen getrennt vorhandene Erbanlagen miteinander zu kombinieren und dadurch vollkommen neue Erbanlagekombinationen zu schaffen. Da die Mendelschen Gesetze nicht nur bei Pflanzen, sondern in gleicher Weise auch bei Tieren gelten, werde ich versuchen, diese Gesetzmäßigkeiten an Beispielen aus der Kaninchenzucht zu erläutern.
Das I. Mendelgesetz ist das Uniformitätsgesetz
Es besagt, dass alle Nachkommen aus der Kreuzung reiner Rassen uniform, d. h. untereinander gleich sind. Nun gibt es bei Kaninchen und anderen Arten, bei denen eine Selbstbefruchtung nicht möglich ist, keine Rassen, die in allen Erbanlagen reinerbig (homozygot) sind. Es genügt aber, sich auf die in reinerbiger Form vorliegenden Erbanlagen, z. B. für Fellfarbe, Haarlänge u. a. zu konzentrieren, um auch aus Kreuzungsexperimenten bei Kaninchen die Mendelschen Gesetze abzuleiten und die zu erwartenden Geno- und Phänotypen im Voraus zu bestimmen.
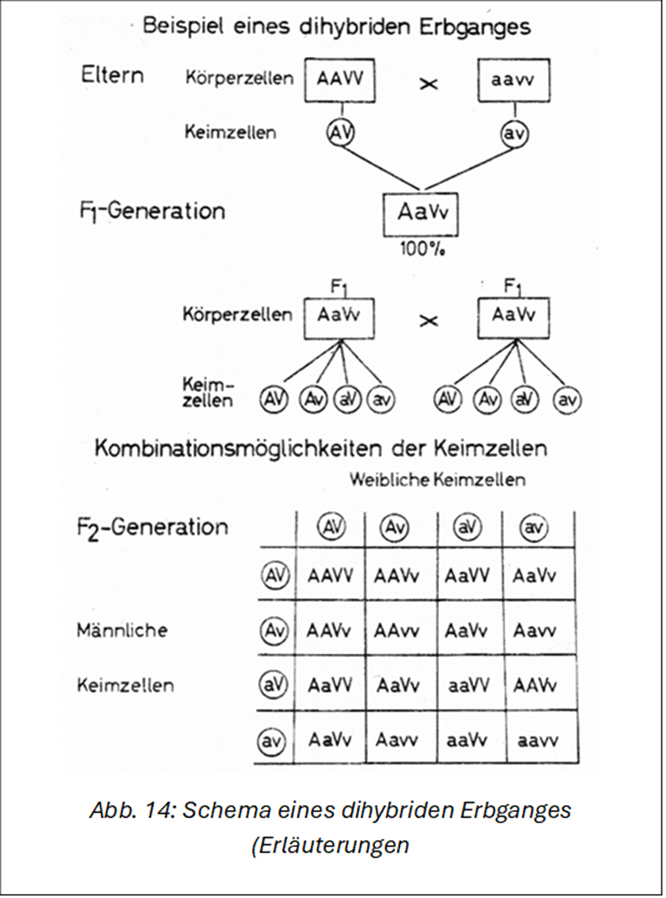
Beispiel: Kreuzt man ein Angorakaninchen (Langhaar) mit einem Weißen Neuseeländer (Normalhaar), so sind alle Nachkommen normalhaarig, unabhängig davon, ob man einen Angorarammler mit einer Weißen Neuseeländerhäsin paart oder die reziproke (wechselseitige) Paarung, nämlich Angorahäsin × Weiße Neuseeländerrammler vornimmt (Abb. 13). Grund: Die Anlage für Normalhaar (V) ist dominant über die von Langhaar (v). Die Ausgangsgeneration wird als P-Generation (Parentes, Eltern), die erste Nachkommengeneration mit F₁ (Filial-, Tochtergeneration) bezeichnet. Bei dem angeführten Beispiel handelt es sich um einen dominanten Erbgang, bei dem nur ein unterschiedliches Merkmal, nämlich die Haarlänge, berücksichtigt wird (monohybrider Erbgang). Aber auch bei Berücksichtigung von 2 oder mehreren unterschiedlichen Genpaaren ist die F₁ einheitlich (Abb. 14).
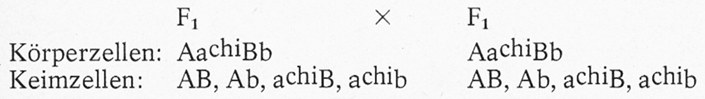
Auf die intermediäre Vererbung bei der Wunderblume mit roten und weißen Blüten wurde bereits hingewiesen. Die F₁ aus einer Kreuzung von weißblühenden und rotblühenden Pflanzen zeigt einheitlich rosafarbige Blüten. Beim Kaninchen sind mir keine vergleichbaren Beispiele bekannt. Es gibt zwar auch hier Erbgänge, z. B. bei Paarungen von Tieren mit unterschiedlicher Größe, Ohrenlänge u. a., bei denen die Merkmalsprägung in der F₁ zwischen der der Ausgangstiere liegt. Da das Entstehen dieser und ähnlicher Merkmale jedoch durch Zusammenwirken vieler, im Einzelnen nicht näher bekannter Erbfaktoren erfolgt, gibt es in der F2 keine klaren Aufspaltungsverhältnisse, wie das bei einfachen Erbvorgängen der Fall ist (vgl. II. Mendelgesetz). Es treten vielmehr zahlreiche Zwischenformen mit fließenden Übergängen in Erscheinung, bei denen sich die meisten entsprechend der Gauß'schen Kurve um den Mittelwert anhäufen, während die extremen Ausgangsformen selten auftreten.
Bei umweltabhängigen Merkmalen, z. B. Körpergröße u. a., kann ihre Ausprägung außerdem durch Umweltfaktoren, wie Fütterung, Haltung, Krankheitserreger, Temperatur u. a., z. T. erheblich modifiziert werden.
Das II. Mendelsche Gesetz ist das Spaltungsgesetz
Wir haben gesehen, dass nach dem I. Mendelschen Gesetz die F₁ aus Kreuzungen reiner Rassen mit unterschiedlichen Merkmalen gleich ist. In unserem Beispiel (Abb. 13) sind alle F₁-Tiere normalhaarig, weil die Anlage für Normalhaar (V) dominant über die Anlage für Langhaar (v) ist. Paart man nun die F₁ untereinander, so tritt nach dem II. Mendelschen Gesetz eine Aufspaltung mit bestimmten Zahlenverhältnissen in Erscheinung. Bei einem monohybriden dominanten Erbgang (unser Beispiel) gleichen 75% der F2 dem dominanten Ausgangspartner (sie sind normalhaarig). 25% sind langhaarig wie der rezessive Ausgangspartner. Das Verhältnis von Normalhaar zu Langhaar ist also 3:1, wie es das II. Mendelsche Gesetz verlangt.
Beim intermediären Erbgang (Wunderblume) ist das Verhältnis rot: rosafarbig: weiß wie 1:2:1. Bei den angegebenen Verhältniszahlen handelt es sich allerdings um Wahrscheinlichkeitswerte, die für den Durchschnitt großer Zahlen gelten. Bei kleineren Nachkommenzahlen, wie sie z. B. innerhalb eines Wurfes oder weniger Würfe auftreten, sind da- gegen z. T. erhebliche Abweichungen möglich. Die hier am Beispiel einer Kreuzung von Angora X Normalhaar erläuterten Verhältnisse gelten für alle monohybriden dominanten Erbgänge. In allen Fällen zeigt die F₁ einheitlich die Wirkung des dominanten Gens. In der F2 ergibt sich ein Aufspaltungsverhältnis von 3:1. Drei F2-Tiere gleichen im Durchschnitt dem dominanten, eines dem rezessiven Ausgangstier.
Von den 3 Tieren mit dem dominanten Merkmal ist allerdings nur 1 reinerbig. Die anderen 2 sind spalterbig. Das rezessive Merkmal kann dagegen nur in reinerbiger Form in Erscheinung treten. Das erleichtert die Züchtung auf rezessive Merkmale, weil man mit Sicherheit weiß, dass derartige Phänotypen, untereinander gepaart, rein weitervererben. In Abb. 13 sind die reinerbigen F2-Tiere mit VV und vv, die spalterbigen mit Vv gekennzeichnet. Darunter sind weitere Paarungsmöglichkeiten innerhalb der F2 und (in der untersten Reihe) die daraus zu erwartenden Nachkommen angegeben. Die Phänotypen sind bildlich dargestellt. Ihr Genotyp ist darunter durch Buchstaben angegeben. Es dürfte dem Leser nicht schwerfallen, die reinerbigen von den spalterbigen Tieren zu unterscheiden.
In Abb. 14 ist ein dihybrider Erbgang dargestellt, bei dem ein farbiges Normalhaarkaninchen mit einem albinotischen Langhaarpartner (Angora) gekreuzt wurde. Anstelle der Tiere sind aus Platzgründen Buchstaben als Gensymbole verwendet worden, über deren Bedeutung bereits gesprochen wurde. Von den in Abb. 14 verwendeten Symbolen bedeutet A: Pigmentierung, V: Normalhaar, a: Albinismus und v: Langhaar. Paart man nun z. B. Alaska oder eine beliebige andere pigmentierte Normalhaarrasse mit Angora, so erhält man eine dihybride (doppeltspalterbige) F₁ mit der abgekürzten Genformel AaVv. Alle F₁-Tiere sind normalhaarig und pigmentiert, weil A dominant ist über a und V über v. Über die Art der Pigmentierung lassen sich bei Kreuzungen mit albinotischen Partnern keine Angaben machen, weil man nicht weiß, welche Farbgene der albinotische Partner besitzt, da diese durch den Albinofaktor nicht zur Wirkung kommen. Wir wollen diese bei albinotischen (rotäugig-weißen) und leuzistischen (blau- äugig-weißen) vorhandenen Besonderheiten zunächst einmal vergessen und uns mit dem Ausdruck „pigmentiert" begnügen.
Paart man nun die F₁-Tiere untereinander (AaVvX AaVv), so entstehen bei der Reduktionsteilung je 4 verschiedene Arten von Keimzellen (Ei- und Samenzellen). Um nun die bei der Befruchtung sich ergebenden Möglichkeiten besser übersehen zu können, benutzt man am besten das unter F2 dargestellte Erbschema. Wir haben dabei die Keimzellen des weiblichen Partners in der waagerechten Zeile (oben) und die des männlichen in der senkrechten (links) eingeordnet. Bei umgekehrter Anordnung käme man zum gleichen Ergebnis. Dabei muss man in beiden Fällen die richtige Reihenfolge einhalten. Kombiniert man nun jede Art der männlichen mit jeder Art der weiblichen Keimzellen, wie das ja auch bei der Befruchtung geschehen kann, so ergeben sich insgesamt 16 Möglichkeiten, von denen allerdings einige identisch sind. Der Leser möge die gleichen Gentypen selbst herausfinden. Insgesamt treten folgende Phänotypen in Erscheinung: 9 normalhaarig-pigmentiert
3 normalhaarig-weiß
3 langhaarig-pigmentiert
1 langhaarig-weiß
Hier ergibt sich also ein Verhältnis von 9:3:3:1.
Von den 9 normalhaarig-pigmentierten Tieren ist 1 reinerbig, 4 sind einfach und 4 doppeltspalterbig. Von den 3 normalhaarig- weißen Nachkommen ist ebenfalls 1 reinerbig und 2 sind einfach- spalterbig. Das gleiche gilt sinngemäß für die langhaarig-pigmentierten Tiere. Das langhaarig-weiße Tier ist reinerbig. Es treten im vorliegenden Beispiel also 2 neue Kombinationstypen in reinerbiger Form in Erscheinung. Es handelt sich um ein pigmentiertes Langhaartier (AAvv) und ein albinotisches Normalhaarkaninchen (aaVV). Wenn man das Schema richtig aufgebaut hat, befinden sich alle reinerbigen Tiere in der Diagonalen von links oben nach rechts unten. Auf weitere Möglichkeiten, die sich z. B. bei Rückkreuzungen oder Paarungen bereits spalterbiger Eltern ergeben, kann hier nicht eingegangen werden. Der Aufbau des Erbschemas wird umso schwieriger, je mehr unterschiedliche Erbanlagen die Eltern aufweisen. Unterscheiden sich die Eltern in drei Erbanlagenpaaren, so ergeben sich bei Paarungen der F₁ untereinander je 8 unterschiedliche Sorten von Keimzellen und 64 Kombinationsmöglichkeiten. Bei 4 unterschiedlichen Genpaaren sind es 16 verschiedene Keimzellensorten und 16 x16 = 256 Kombinationsmöglichkeiten usw. Die maximal mögliche Zahl unterschiedlicher Keimzellen und befruchteter Eizellen ist von der Anzahl der Chromosomenpaare abhängig. Da beim Kaninchen 44 = 22 Paar Chromosomen vorhanden sind, ergeben sich 222, das sind über 8 Millionen Möglichkeiten der Bildung unterschiedlicher Keimzellen und über 70 Billionen unterschiedlicher Zygoten. Welche dieser Möglichkeiten bei den relativ wenigen Nachkommen zum Tragen kommen, bleibt dem Zufall über- lassen.

Abb. 15: Vom Verfasser aus Roten Neuseeländern und Chinchilla gezüchtetes Schwarzgrannen-Kaninchen.
In diesem Zahlenspiel, das die Vielfältigkeit der Natur verständlich machen soll, sind nur die erblich bedingten Variationsmöglichkeiten berücksichtigt worden. Hinzu kommen noch die Einflüsse der Umwelt (Ernährung, Haltung, Krankheitserreger u. a.), welche die Wirkungen der Erbanlagen modifizieren, d. h. phänotypisch, nicht aber erblich verändern.
Der Züchter braucht trotz dieser großen Vielfältigkeiten jedoch nicht zu verzweifeln. Die meisten unterschiedlichen Genkombinationen machen sich phänotypisch kaum bemerkbar. Ferner treten alle Genpaare, die beide Eltern in reinerbiger Form besitzen, auch reinerbig bei allen Nachkommen wieder in Erscheinung. Je mehr Erbanlagenpaare also in reinerbiger Form vorhanden sind, desto ausgeglichener werden auch die Nachkommen sein. Es gibt – sehr selten – auch Fälle mit genetischer Identität, d. h. völliger erblicher Gleichheit. Das ist z. B. bei eineiigen Zwillingen der Fall. Auch sog. „Klone“, die hauptsächlich bei Pflanzen durch ungeschlechtliche Fortpflanzung entstehen, z. B. Ableger, Kartoffelknollen u. a., sind erblich völlig gleich. In der letzten Zeit ist die Entwicklung von Klonen auch bei Tieren, und zwar bei Fröschen geglückt. Sie wird von Fachwissenschaftlern im Prinzip auch bei anderen Tierarten als machbar angesehen. Dabei werden die Kerne aus den Eizellen herausoperiert und durch diploide Körperzellen ersetzt. Es bedarf aber noch weiterer Erfahrungen und einer verbesserten Technik, um die Erfolgschancen weiter zu erhöhen.
Das III. Mendelsche Gesetz:
Unabhängigkeit und Möglichkeit der Neukombination der Gene
Einiges zu diesem Thema Gehörende wurde im Vorhergehenden bereits angesprochen. Neukombinationen von Erbanlagen, die zunächst getrennt bei verschiedenen Tieren vorhanden waren, bilden die Grundlage der Züchtung. Alle Kaninchenrassen sind durch Kombinationszüchtung entstanden.
In Abb. 14 wurde bereits eine einfache Kombinationszüchtung dargestellt. Bei Kreuzungen von pigmentierten Normalhaarkaninchen mit albinotischen Langhaartieren konnten sowohl der Langhaarfaktor (v) mit dem Grundfaktor für Pigmentierung (A) als auch die Anlage für Normalhaar (V) mit dem Albinofaktor (a) in reinerbiger Form kombiniert werden.
Interessanter ist die Entwicklung des von mir gezüchteten Schwarzgrannenkaninchens (Abb. 15), bei dem rezessive Gene miteinander kombiniert wurden (Niehaus, 1969). Das hat gegenüber dominant-rezessiven Kreuzungen den Vorteil, dass die doppeltrezessiven Endprodukte eindeutig erkannt und rein weitergezüchtet werden können.
Als Ausgangsrassen für die Züchtung von Tieren mit weißem Haarkleid und braunen Augen wurden Rote Neuseeländer mit Großchinchilla gekreuzt. Die F₁-Tiere waren alle – wie erwartet – hasenfarbig.
Lassen wir bei der Weiterpaarung der F₁-Bastarde die y-Faktoren unberücksichtigt, weil sie auf das Aussehen des Endproduktes keinen Einfluss haben, so ergibt die Weiterpaarung folgendes Bild:
Fasst man die im Schema angegebenen Typen nach ihrer Erscheinungsform zusammen, so ergeben sich 9 hasenfarbige
3 chinchillafarbige
3 rötlich-gelbe Tiere
1 Tier mit weißer Fellfarbe
Dabei sind die in der verkürzten Erbformel nicht angegebenen y-Faktoren sowie die Anlage für Farbverteilung (G) nicht berücksichtigt worden. Wir haben hier, wie in Abb. 14, wieder ein Verhältnis der Phänotypen von 9:3:3:1 entsprechend der von Mendel im Jahr 1866 für einen dihybriden Erbgang aufgestellten Regel.
Wie in Abb. 14 sind auch hier alle in der Diagonalen von links oben nach rechts unten aufgeführten Genotypen reinerbig in den aufgezeichneten Genpaaren. Dabei ist ein sehr interessanter neuer Kombinationstyp (achiachibb) entstanden. Nachtsheim (1977) hatte bereits 1949 darauf hingewiesen, dass bei einer reinerbigen Kombination des Chinchillafaktors achi mit der Anlage für gelbe Farbe (b) Tiere mit weißer Fellfarbe entstehen. „Kommen nun Chinchilla- und Gelbfaktor zusammen, so unterdrückt der Chinchillafaktor das gelbe Pigment, das der Gelbfaktor allein übrig lässt, und das Tier wird weiß.“ Die dunklen Grannenspitzen wider- sprechen den Angaben von Nachtsheim nicht, weil es nicht selten vorkommt, dass die Farbe der Grannenspitzen von der übrigen Fellfarbe abweicht. Ferner ist es uns gelungen, die dunklen Grannen durch fortlaufende Selektion wegzuzüchten und dadurch Tiere mit reinweißem Fell und dunkelbrauner Augenfarbe zu erhalten.
Es sei noch bemerkt, dass nach noch nicht abgeschlossenen Experimenten von Schmitt (1978) die Anwesenheit des Farbverteilungsfaktors G vermutlich einen wichtigen Einfluss auf das Erscheinungsbild dieses Genotyps ausübt. Ich darf annehmen, dass Schmitt zu gegebener Zeit die Ergebnisse seiner Experimente veröffentlichen wird. Im Übrigen möchte ich es dem Leser überlassen, aus dem Aufspaltungsschema neben den reinerbigen Genotypen auch diejenigen herauszufinden, die einfach- oder doppeltspalterbig sind. Die Züchtungsverfahren werden umso komplizierter, je mehr Gene miteinander kombiniert werden sollen. So ergeben sich bei einem trihybriden Erbgang je 8 verschiedene Arten von Keimzellen und 64 Kombinationsmöglichkeiten. Als Beispiel für einen trihybriden Erbgang sei die Züchtung eines leuzistischen (blau- äugigen) Kaninchens aus Weißen Wienern und Angorakaninchen erwähnt (Niehaus, 1954).
Erfahrene Züchter wissen, dass das Zuchtziel durch die Kombination von zwei oder mehreren Genpaaren noch lange nicht erreicht ist. Es kommt dann die mühselige Feinarbeit zur Verbesserung der Größe, der Körperform, des Felles, der Gesundheit, der Fruchtbarkeit und anderer Merkmale, die durch das Zusammenwirken vieler, im Einzelnen nicht näher bekannter Anlagen bedingt und durch die Mendelgesetze nicht zu erfassen sind. Da die von den einzelnen Züchtern gehaltenen Tierzahlen für populationsgenetische Züchtungsverfahren zu klein sind, muss der Züchter versuchen, durch fortlaufende Auslese und Verpaarung der dem Zuchtziel am nächsten kommenden Tiere eine Verbesserung in dem gewünschten Sinne zu erreichen. Dass man auch damit Erfolge erzielen kann, beweist der hohe Standard der Rassekaninchenzucht.
Dieser Artikel ist in erster Linie für genetisch interessierte Kaninchenzüchter geschrieben worden. Ich habe mich deshalb auch bemüht, die Vererbungsvorgänge möglichst an Beispielen aus der Kaninchenzucht zu erläutern. Parallelen dazu sind aber auch bei anderen Tierarten (Rinder, Mäuse, Hunde, Katzen, Hühner u. a.) bekannt. Ich hoffe, dass Hinweise auf moderne Erkenntnisse der Vererbungsvorgänge den Leser nicht allzu sehr verwirren. Leider war eine ausführlichere Darstellung im Rahmen dieser Arbeit nicht möglich. Auch auf dem Gebiete der einfachen Erbvorgänge konnte nur eine Auswahl, der mir wichtig erscheinenden Fakten und Zusammenhänge getroffen werden.
Mögen diese Ausführungen aber dazu beitragen, das Interesse der Züchter für die wunderbaren Vorgänge im Reiche des Lebendigen zu wecken und Denkanstöße für Diskussionen in Züchterkreisen zu geben.
Literatur
v. Ditfurth, H.: Was ist Leben? X-Magazin für Naturwissenschaft und Technik, Heft 8, Deutsche Verlags-Anstalt, Stuttgart, 1972
Kühn, A.: Grundriss der Vererbungslehre, Quelle und Meyer, Heidelberg, 1973 Nigon, V. u. Lueken, W.: Allgemeine Biologie, Band 4, Vererbung, Gustav-Fischer- Verlag, Stuttgart, 1976
Nachtsheim, H.: Vom Wildtier zum Haustier, 3. Aufl., Paul Parey-Verlag, Berlin und Hamburg, 1977 Gleichauf, R.: Züchtungs- und Vererbungslehre für Geflügelzüchter, Fritz Pfenningstorff, Berlin und Stuttgart, 1972
Dietrich, G., Stöcker, F. et al.: Fachlexikon abc Biologie, Harri Deutsch, Thun und Frankfurt/M., 1976 Geissler, Libbert, Nitschmann, Petersein et al.: Kleine Enzyklopädie „Leben", VEB Bibliographisches Institut, Leipzig, 1978
Niehaus, H.: Züchtung des Schwarzgrannen-Kaninchens, Das Blaue Kaninchen-Jahrbuch, Oertel+Spörer, Reutlingen, 1969 Schmitt, H.: Schriftliche Mitteilungen,
Niehaus, H.: Neuzüchtung eines leuzistischen (blauäugigen) Angorakaninchens. Dt. Kleintier-Züchter, Ausg. Kaninchen, 2, Oertel + Spörer, Reutlingen, 1954