

Heinrich Niehaus, Celle
Institut für Kleintierzucht der Forschungsanstalt für Landwirtschaft
Braunschweig-Völkenrode
„Das Blaue Jahrbuch“ 1975
Einleitung
Unsere Kenntnisse auf den verschiedenen Gebieten der Kaninchenzucht und -haltung, u. a. auch der Verdauungsvorgänge, sind noch lückenhaft und ungenau. Das ist an sich verwunderlich, weil das Kaninchen schon seit langem als Versuchstier sowie für die Produktion von Kaninchenfleisch, Angorawolle und Kaninchenfellen eine große Bedeutung besitzt.
Ich werde im Folgenden versuchen, die mir aus der Literatur bekannten Daten über die Verdauungsvorgänge beim Kaninchen und die dabei auftretenden Besonderheiten in den mir wesentlich und interessant erscheinenden Punkten aufzuzeigen und die Literaturangaben durch eigene Versuchsergebnisse und Beobachtungen zu ergänzen.
Zweck der Verdauung
Die vom Kaninchen aufgenommenen Futterstoffe müssen zu- nächst bestimmte Veränderungen erfahren, bevor die verwertbaren Teile in den Blutkreislauf gelangen und zur Ernährung der Körperzellen, zum Aufbau von Fleisch, Fett, Knochen, Drüsen, Fermenten, Hormonen u. a. oder zum Energiestoffwechsel (Erhaltung der Körperwärme, für die Leistungswärme oder Bewegungsenergie) verwendet werden können.
Den Aufschluss der im Futter enthaltenen Nahrungsstoffe durch mechanische und chemische Einwirkungen nennt man Verdauung. Die mechanischen Vorgänge bestehen in der Zerkleinerung, Einspeichelung, Durchmischung und Fortbewegung des Futters. Durch chemische (fermentative) Einwirkungen werden wasserunlösliche oder schwerlösliche hochmolekulare organische Nährstoffe (Kohlehydrate, Eiweiß und Fett) zu niedermolekularen wasserlöslichen Verbindungen abgebaut. Wasser, Vitamine und die meisten Salze bedürfen keiner derartigen Veränderung, um in die Blutbahn zu gelangen. Die nicht verdauten Futterbestandteile werden als Kot (Faeces) ausgeschieden.
Die Verdauungsvorgänge erfolgen im Verdauungstrakt, einem langen Schlauch, der aus mehreren Abschnitten besteht und mit kleineren und größeren Ausbuchtungen versehen ist. Er beginnt am Maul und endet am After. Die wesentlichen Teile des Verdauungstraktes sind:
1. Mundhöhle
2. Magen
3. Dünndarm mit den Anhangdrüsen Leber und Bauchspeicheldrüse
a) Zwölffingerdarm (Duodenum)
b) Leerdarm (Jejunum)
c) Hüft- oder Krummdarm (Ileum)
4. Dickdarm
a) Blinddarm (Caecum oder Coecum)
b) Grimmdarm (Colon)
c) Mastdarm (Rectum)
Magen und Darm eines 4 Monate alten Kaninchens (WN) sind auf Abb. 1 zu erkennen.
Wie das Foto zeigt, besitzen Kaninchen einen langen und geräumigen Verdauungstrakt, in dem sie größere Futtermengen bewältigen können, wie das zur Deckung des Nahrungsbedarfs in der Wildbahn und bei der herkömmlichen Fütterung der Hauskaninchen erforderlich ist. Es hat sich gezeigt, dass Kaninchen auch an eine Fütterung mit konzentrierten Allein-Fertigfuttermischungen und Wasser angepasst werden können. Der Prozess einer völligen Anpassung dauert meist viele Generationen und erfordert eine fortlaufende Selektion der am besten angepassten Tiere. Die vorliegenden Versuchsergebnisse reichen noch nicht aus, um sich ein ausreichendes Urteil über die auf diesem Gebiet vorhandenen Möglichkeiten machen zu können. Bei einer übergangslosen Umstellung auf Alleinfutter treten zunächst häufig Fruchtbarkeitsstörungen und Verfettung bei Alttieren sowie Durchfall bei Jungtieren auf. Bei Umstellung der Haltung von Streu auf Drahtgitterböden sind die Tiere in den ersten Generationen anfällig gegen „wunde Läufe“.
Länge und Inhaltsgewichte der Verdauungsorgane
Nach Mangold (1950) besitzt der Darm des Kaninchens eine Länge von etwa 4-6 m, was das 9- bis 11fache der Körperlänge ausmacht. Kametaka (1973) ermittelte eine Darmlänge von 5,8 (4,7-6,5) m, entsprechend einer Relation zur Körperlänge von etwa 10:1. Hinsichtlich der Kapazität der Verdauungsorgane weichen die Angaben der älteren Literatur (Mangold, 1950, Jung, 1962 u. a.) z. T. erheblich von denen von Kametaka (1973) veröffentlichten Daten ab.
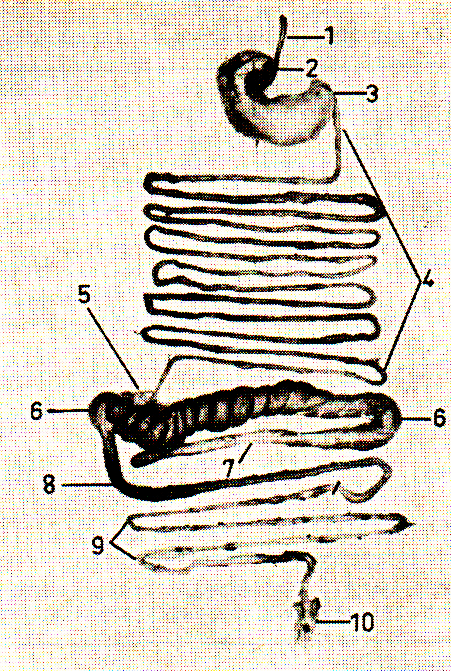
Abb. 1: Der Verdauungsapparat eines zwölf Wochen alten Weißen Neuseeländer-Kaninchens. Die Verdauungsorgane wurden zum Zwecke einer besseren Übersicht aus ihrer natürlichen Lage herausgelöst. Es bedeuten: 1: Speiseröhre, 2: Mageneingang, 3: Magenausgang, 4: Dünndarm, 5: Sacculus rutundus, 6: Blinddarm, 7: Blinddarmfortsatz, 8: Grimmdarm, 9: Mastdarm u. 10: After.
Während von Mangold, Jung u. a. das Fassungsvermögen des Kaninchenmagens mit durchschnittlich 50 ccm bzw. 50 ml angegeben wird, fand Kametaka (1973) Inhaltsgewichte von g. Beim Blinddarm liegen die Verhältnisse umgekehrt. Von Mangold (1950), Dorn (1973) u. a. wird der Blinddarm als sehr geräumig bezeichnet, der einen großen Teil der Bauchhöhle anfüllt. Nach Jung (1962) fasst der Blinddarm das 8- bis 12fache des Magens. Kametaka (1973) ermittelte Inhaltsgewichte von g. Daraus errechnet sich ein Verhältnis von Magen- zu Blinddarminhalt von nur 1:1,2.
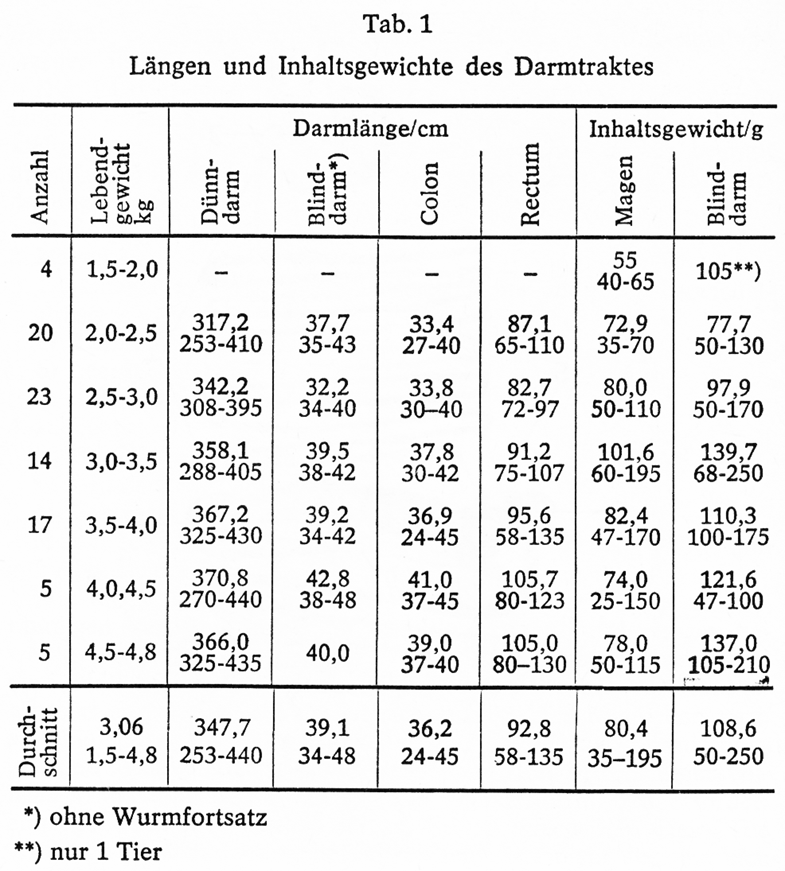
Die ersten Teilergebnisse eigener, noch nicht abgeschlossener Untersuchungen über Darmlänge und Inhaltsgewichte bei Kaninchen mittelschwerer Rassen (WN, Kal, WRex) und Kreuzungen sind in Tab. 1 zusammengestellt. Es sei noch bemerkt, dass die Tiere und ihre Vorfahren nur Alleinfutter und Wasser zur freien Aufnahme erhielten und ohne vorhergehende Nüchterung geschlachtet wurden, dass es sich ferner überwiegend um noch im Wachstum befindliche Jungtiere unterschiedlicher Gewichts- und Altersgruppen handelte.
Aus den Zahlen der Tab. 1 ergeben sich folgende Gesamt- und Verhältniswerte:
Darmlänge
Gesamtlänge 515,8 (369-668) cm = 100%
Dünndarm 347,1 (253-440) cm 67,7%
Dickdarm 168,1 (116-228) cm = 32,6%
Demnach entfallen etwa 2/3 der gesamten Darmlänge in Übereinstimmung mit Mangold (1950) auf den Dünndarm und etwa 1/3auf den Dickdarm.
Inhaltsgewichte von Magen und Blinddarm
Magen: 80,4 (35*-195) g
Blinddarm: 108,6 (50*-250) g
Das Verhältnis von Magen- zu Blinddarminhalt beträgt bei unseren Untersuchungen im Durchschnitt 1:1,35. Aus den von Kametaka (1973) veröffentlichten Zahlen ergibt sich ein Verhältnis von 1:1,2. Das entspricht in etwa unseren Ermittlungen, weicht aber erheblich von den von Jung (1962) gemachten Angaben ab, nach denen der Blinddarm 8 bis 12mal so groß ist wie der Magen.
Insgesamt gesehen, waren die Streuungen sowohl der Darmlänge als auch – in besonderem Maße – der Inhaltsgewichte von Magen und Blinddarm bei unseren Untersuchungen sehr groß. Folgende Gründe erscheinen mir hierfür wesentlich:
1. Länge und maximale Kapazität der Verdauungsorgane stehen im Zusammenhang mit der Größe der Tiere. Das ist aus den Zahlen der Tab. 1 deutlich zu erkennen.
*) Die geringsten von uns gefundenen Inhaltsgewichte des Magens (35 g) und des Blinddarms (50 g) stammen von Tieren, denen das Futter 40 Stunden vor dem Schlachten entzogen wurde. Wasser stand ihnen zur freien Verfügung. Die Mägen dieser Tiere waren stark geschrumpft und das verkleinerte Lumen weitgehend mit einem dünnflüssigen Brei gefüllt.
2. Es gibt aber auch bei Tieren gleicher Größe individuell bedingte, z. T. erhebliche Streuungen.
3. Es dürfte ferner nicht bezweifelt werden, dass die Menge des in den Tagen vor der Untersuchung aufgenommenen Futters die Magen- und Darminhalte beeinflusst. Dabei ist zu erwähnen, dass die Wände von Magen und Blinddarm sehr dehnbar sind und ihr Volumen sich dem Inhalt weitgehend angleicht. Das maximale Fassungsvermögen kann deshalb nur bei prallgefülltem Magen und Blinddarm mit ausreichender Genauigkeit ermittelt werden.
4. Die vielfach in Züchterkreisen verbreitete Meinung, die Entwicklung des Verdauungstraktes würde durch die Zusammensetzung des Futters (z. B. ballastreich oder konzentriert) beeinflusst, konnte durch Versuche von Haesler (1930) an Schweinen und Ratten sowie von Lenkeit (1934) an Hühnern nicht bestätigt werden. Mangold (1951) hält die Möglichkeit, die individuellen Schwankungen der absoluten und damit der relativen Darmlänge bei einer Tierart im Wesentlichen auf Einflüsse individuell verschiedener Ernährung zurückzuführen, für widerlegt. Diese Einflüsse können sich nur sekundär in geringem Maße auswirken. Die entwicklungsgeschichtliche Form- und Größengestaltung ist nach Mangold (1951) offenbar erblich bedingt.
5. Allgemein gültige Angaben über Länge und Fassungsvermögen des Verdauungstraktes bei Kaninchen sind für die breitgefächerte Skala der zahlreichen Rassen von den Zwergen bis zu den Riesen nicht zu erstatten. Ich halte es aber für möglich, brauchbare Faustzahlen für eine bestimmte Größenklasse und Korrektionsfaktoren für alle abweichenden Größen zu ermitteln.
Vorgänge im Maul des Kaninchens
Das Futter wird mit Hilfe der Lippen, Zunge und Zähne aufgenommen, durch die Kauwerkzeuge, speziell durch die mahlenden Bewegungen der Backenzähne sorgfältig zerkleinert und durch den schleimhaltigen Speichel in einen schlüpfrigen Futterbrei verwandelt, bevor es abgeschluckt wird und durch die eigene Schwerkraft sowie mit Hilfe peristaltischer Bewegungen der Speiseröhre (Ösophagus) in den Magen gelangt. Der Kauvorgang hat folgende Bedeutung: Beim Kauen werden pflanzliche Zellwände (z. B. beim Grünfutter) zerstört, so dass der Zellinhalt von den im Maul, Magen und Darm abgesonderten Verdauungssäften erreicht und verdaut werden kann. Ferner wird die Gesamtoberfläche der Futterteilchen vergrößert. Da die Verdauungssäfte an der Oberfläche der Futterteilchen angreifen, wird die Verdaulichkeit der Nährstoffe sowohl durch die Zerstörung von Pflanzenzellen als auch durch die Zerkleinerung des Futters erhöht. Die Vermischung des Futters mit größeren Mengen schleimigen Speichels beim Kauen erleichtert das Abschlucken und die Weiterbeförderung des Speichelbreies, bewirkt ein Aufquellen der Kolloide und trägt dadurch ebenfalls zur Verdaulichkeit durch Bakterien und körpereigene Fermente bei („Gut gekaut ist halb verdaut").
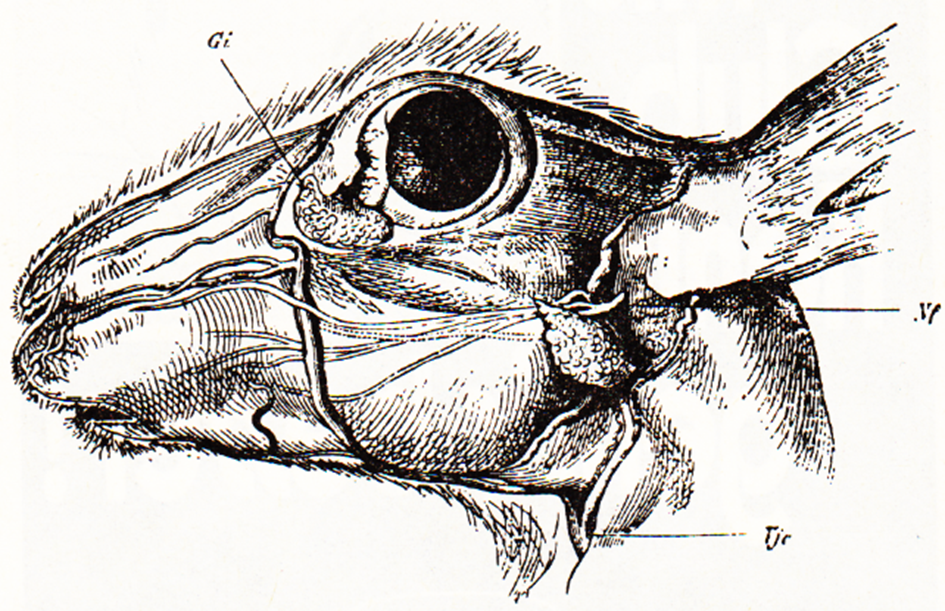
Abb. 2: Der Kopf des Kaninchens mit Ohrspeicheldrüse und Gi Infraorbitaldrüse, Nf Gesichtsnerven, Tjc Halsvene (nach Krause aus Mangold, 1950).
Der gemischte Mundspeichel wird von den beiderseitigen Speicheldrüsen: Ohrspeicheldrüse (Parotis), Unterkieferdrüse (Submaxillaris), Unterzungendrüse (Sublingualis) und der unter dem Auge befindlichen Infraorbitaldrüse (bei anderen Tierarten, z. B. Pferd und Schwein, nicht vorhanden) sowie den Drüsen der Zunge und des weichen Gaumens abgesondert.
Der Speichel besteht zu über 99 Prozent aus Wasser. Er enthält ferner Schleimstoffe (Mucine) sowie Chloride, Phosphate und Bicarbonate von Alkalien und Erdalkalien. Letztere bewirken eine schwachalkalische Reaktion.
Die Menge ihrer Ausscheidungen wird durch die Beschaffenheit des Futters beeinflusst.
Die Sekretion der Speicheldrüsen wird durch vom Nervensystem ausgehende Reize ausgelöst. Eine Aktivierung der zuständigen Nerven erfolgt einerseits durch mechanische und chemische Einwirkungen des Futters auf die Mundhöhlenschleimhaut bei der Futteraufnahme und beim Kauen. Andererseits können – besonders bei hungrigen Tieren – Anblick und Geruch des Futters psychisch (seelisch) bedingten Erregungen des Nervensystems und dadurch eine Sekretion (Absonderung) des Speichels bewirken („Da läuft einem das Wasser [Speichel] im Munde zusammen").
Das Gebiss des Kaninchens (Abb. 3) besteht normalerweise aus 28 Zähnen.
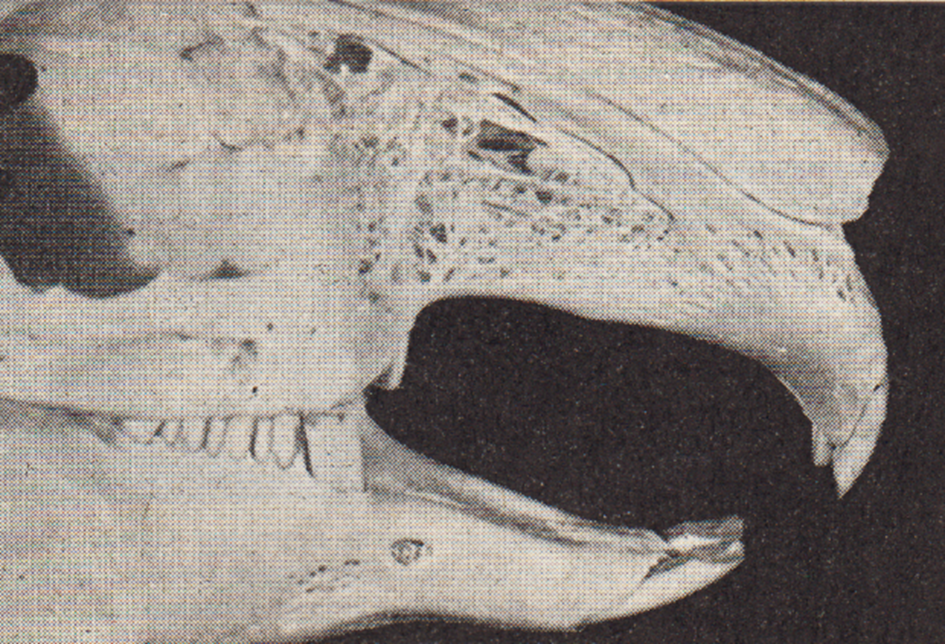
Abb. 3: Normales Gebiss eines erwachsenen Kaninchens mit der typischen Zahnlücke im Ober- und Unterkiefer. Die im Oberkiefer hinter den eigentlichen Schneidezähnen befindlichen verkümmerten Stiftzähne haben dieser Tierordnung den Namen „Doppelzähnige" (Dublicidentata) gegeben.
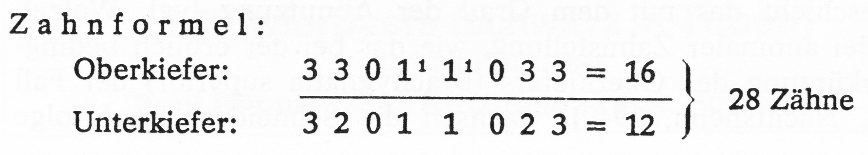
Wie aus der Zahnformel ersichtlich ist, befinden sich im Oberkiefer 16, und zwar 4 (2 Paar) Schneidezähne (Incisivi) und 12 abgeschrägte Backenzähne (6 Prämolaren und 6 Molaren), während der Unterkiefer nur 12 (2 Schneidezähne und 10 Backenzähne) enthält. Zwischen den Schneide- und Backenzähnen befindet sich eine auch bei Nagetieren vorhandene Zahnlücke.
Typisch für Lagomorphen (Kaninchen, Hasen und Pfeifhasen) sind ferner die doppelt angelegten Schneidezähne. Hinter den eigentlichen normal ausgebildeten Schneidezähnen befinden sich im Oberkiefer 2 verkümmerte (rudimentäre) Stiftzähne, die gelegentlich fehlen können. In der Zahnformel sind sie durch die bei- den hochgestellten Zahlen symbolisiert.
Die doppelt vorhandenen Schneidezähne im Oberkiefer der Lagomorpha haben ihnen den Namen „Doppelzähnige“ (Duplicidentata) gegeben. Die Lagomorpha unterscheiden sich u. a. auch durch diese Besonderheit von den eigentlichen Nagetieren (Rodentia), den „Einfachzähnigen“ (Simplicidentata), zu denen Maus, Ratte, Bieber und viele andere Arten gehören.
Abb. 4: Der Unterschied zwischen einem normalen Gebiss (linkes Bild) und einem Gebiss mit verkürztem Oberkiefer (rechtes Bild) ist deutlich zu erkennen. Wenn die Zähne keinen Gegendruck haben, wachsen sie hemmungslos weiter und krümmen sich maulwärts. Das gilt sowohl für die eigentlichen Schneidezähne als auch für die Stiftzähne.
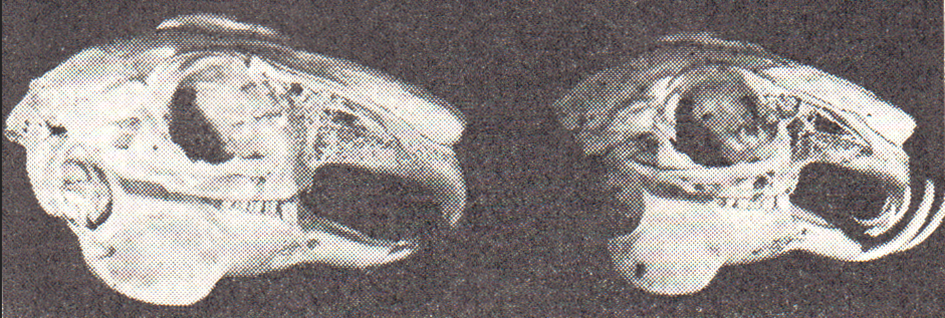
Die charakteristischen Schneidezähne besitzen ein offenes Wurzelende (Hochstrasser, 1972). Sie haben keine Zahnwurzel und sind in der Lage, fortlaufend zu wachsen. Bei normaler Zahnstellung geschieht das mit dem Grad der Abnutzung (vgl. Wetzel, 1931). Bei anomaler Zahnstellung, wie das bei der erblich bedingten Verkürzung des Oberkiefers (Brachygnatia superior) der Fall ist (vgl. Nachtsheim, 1949), wachsen die Schneidezähne infolge 96 mangelnden Gegendruckes ungehemmt weiter, krümmen sich maulwärts und verhindern in zunehmendem Maße die Futteraufnahme (Abb. 4, rechtes Bild). Die befallenen Tiere können schließlich bei voller Futterkrippe verhungern, wenn der Züchter dieses Übel nicht durch wiederholtes Abkneifen der fortlaufend wachsenden Schneidezähne verringert.
Derartige Tiere bleiben trotz aller Bemühungen des Züchters meist in der Entwicklung zurück. Für die Zucht sind sie natürlich nicht geeignet, weil die rezessive (überdeckbare) Anlage auf die Nachkommen vererbt wird. Es sei noch bemerkt, dass der Grad der Kiefermissbildung biologisch bedingte Streuungen aufweist, so dass Tiere mit geringfügigen Missbildungen auch ohne Hilfe des Züchters überleben können. Für die Zucht sollte man aber solche Tiere nicht einsetzen.
Falsch ist die gelegentlich von Züchtern und auch in der Literatur vertretene Meinung, „lange Zähne“ wären auf mangelnde Nagemöglichkeiten zurückzuführen. Trotz langjähriger Haltung eigener Tiere in Drahtkäfigen ohne Nagemöglichkeiten und Fütterung mit gekörntem Alleinfutter und Wasser konnte kein vermehrtes Auftreten anomal langer Zähne beobachtet werden. Bei den von uns untersuchten Fällen war das Übel stets mit einer Verkürzung des Oberkiefers verbunden.
Die Schneidezähne des Kaninchens sind immer meißelscharf. Sie werden durch Benutzung nicht stumpf wie Meißel, Beil und Axt, sondern schleifen sich fortlaufend dadurch, dass die Vorderseite der unteren Schneidezähne an der Hinterfläche der oberen entlanggleitet.
Dabei werden auch die Stiftzähne abgenutzt, die bei anomaler Zahnstellung weiterwachsen, wie die eigentlichen Schneidezähne (vgl. Abb. 4). Dass auch die Schneidezähne des Unterkiefers scharf bleiben, erklärt Hochstrasser (1972) durch gelegentliches Vorbeißen, bei dem die oberen Schneidezähne die unteren abschleifen.
Der Schleifeffekt wird noch dadurch verstärkt, dass die harte Schmelzschicht, die den ganzen Zahn des Kaninchens umgibt, an der Rückseite wesentlich dünner ist als an der Vorderseite.
Kaninchen scheiden mit dem Speichel ein diastatisches Ferment (Ptyalin) aus, das den Nährstoff Stärke bis zu Malzzucker (Maltose) aufspaltet. Stärke ist in hohem Prozentsatz in Getreidekörnern, Kartoffeln u. a. enthalten. Diastase kommt nach Nehring (1952) in größeren Mengen nur im Speichel von Menschen und Schweinen vor. Sie wurde aber auch im Speichel von Kaninchen, Affen, Ratten, Mäusen, Meerschweinchen, Igeln, Eichhörnchen und Hamstern gefunden (Hoppe-Seyler/Thierfelder, 1953). Der Speichel von Hund, Katze, Pferd, Rind u. a. enthält dagegen kein diastatisches Ferment.
Diastase hat seine größte Wirksamkeit bei schwach alkalischer Reaktion, wie sie im Speichel vorhanden ist. Die Wirkung wird gebremst, wenn das Futter in den Bereich der Magensalzsäure gelangt und die Reaktion sauer wird. Andere Fermente spielen nach Mangold (1950) demgegenüber bei der Mundverdauung nur eine untergeordnete Rolle. Insgesamt gesehen finden im Maul des Kaninchens in erster Linie eine mechanische Zerkleinerung, Einspeichelung und ein Aufquellen des Futters und nur in geringem Umfang eine chemische Verdauung statt.
Abb. 5: Prallgefüllter Magen des Kaninchens (nach Jung, 1962
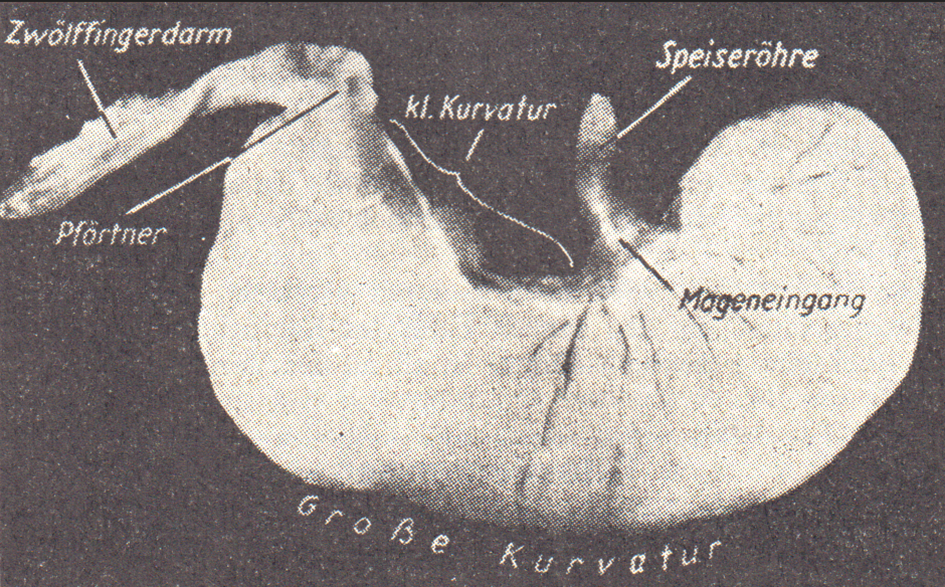
Der Magen und seine Funktionen
Der einhöhlige Magen des Kaninchens (Abb. 5, 6 und 7) dient wie bei anderen monogastrischen, d. h. mit einhöhligen Mägen versehenen Tieren in erster Linie der Speicherung und langsamen Abgabe des Nahrungsbreies in den Dünndarm, wo der Hauptteil der chemischen Verdauung stattfindet. Die wichtigsten Teile des häutigen Magens sind: 1. Mageneingang mit dem fest verschließbaren Cardiamuskel,
2. der große blindsackartige Fundusteil (rechts),
3. die Pylorusregion mit dem erweiterten Pförtnervorhof und dem Magenausgang (links).
Der gesamte Magen des Kaninchens ist mit einer drüsigen Schleimhaut versehen, die an der Bildung des Magensaftes (Salzsäure, Pepsin und Lab) beteiligt ist.
Die Muskulatur des Kaninchenmagens ist mit Ausnahme der Pylorusregion (am Magenausgang) nur schwach ausgebildet. Infolge geringer Magenperistaltik lagert sich das aufgenommene Futter schichtweise ab.
Es wird zunächst im Wesentlichen durch seine Schwerkraft, ferner – bei prallgefülltem Magen – zusätzlich durch die Spannung der Magenwand in Verbindung mit der schwachen Magenperistaltik in Richtung Magenausgang bewegt. Dort wird es von der starken Pylorusmuskulatur erfasst, durchmischt und in den Dünndarm befördert.
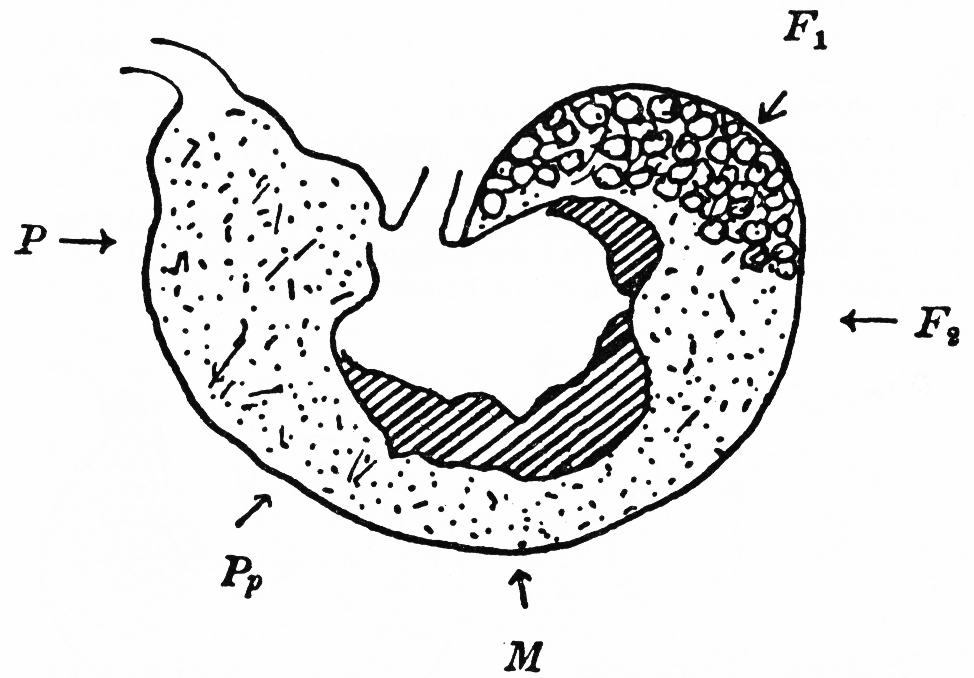
Abb. 6: Längsschnitt durch einen Kaninchenmagen. Schichtung des Mageninhalts. Rechts im Fundusteil (F1) Blinddarmkot, links der Pylorusteil mit erweitertem Pförtnervorhof (Atrum pylori). (Nach Grützner aus Mangold, 1950).
Die starken Muskelstränge der Pylorusregion können bei geschlachteten Tieren deutlich erkannt werden.
Bei eigenen Schlachtungen wurden meist prallgefüllte, niemals leere Mägen gefunden (vgl. Dorn, 1972, Mangold, 1950, Werner, 1969, Jung, 1962). Wurde den Tieren vor dem Schlachten das Futter für 1 oder 2 Tage entzogen – Wasser stand ihnen zur beliebigen Aufnahme zur Verfügung -, so konnte eine z. T. erhebliche Schrumpfung des Magens festgestellt werden. Derartige Mägen waren mit einem dünnen Futterbrei gefüllt. Ferner wurden fast immer Haare in unterschiedlicher Konzentration gefunden, vielfach in Form verfilzter Haarstränge.
Blinddarmkot wurde in geringer Menge bei geschrumpften, in weit größerem Umfange in gefüllten Mägen gefunden. Dieser Blinddarmkot befand sich bei unseren Untersuchungen immer im oberen Teil des Fundus (vgl. Abb. 6). Um ganz sicherzugehen, dass es sich bei den geformten Elementen tatsächlich um Blinddarmkot handelte, haben wir einige Tiere durch Anlegen eines Kragens und Fixierung der Tiere an der Aufnahme von Blinddarmkot gehindert. Wie erwartet, enthielten deren Mägen keine geformten Elemente.
Die von Dorn (1973) angegebene Formung des Mageninhaltes zu kleinen Kügelchen im Bereich des Magenausganges (Abb. 7) konnte bei unseren Untersuchungen in keinem Fall festgestellt werden.
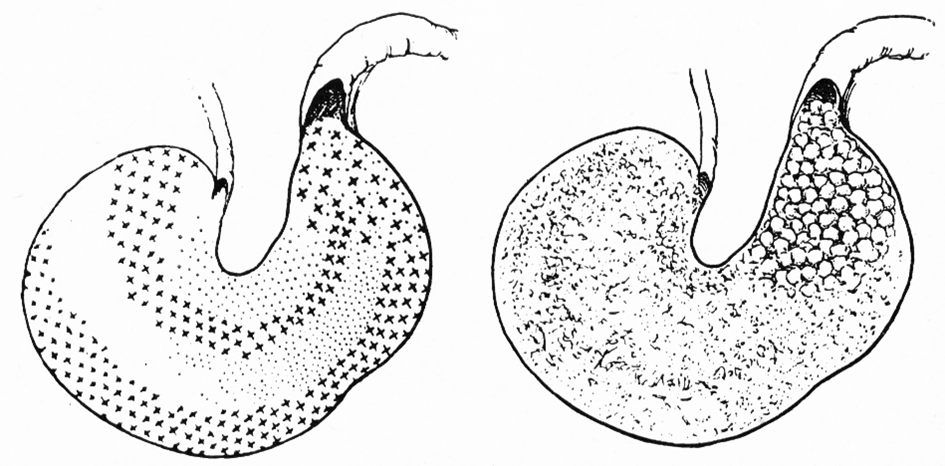
Abb. 7: (Nach Dorn 1973). Bemerkung des Autors: Linke Abb.: „Die Schichtung des Speisebreis im Magen. Rechte Abb.: Die Durchmischung des Mageninhaltes ist erst im Bereich des Magenausganges erfolgt. Das aufgenommene Futter wird hier zu kleinen Kügelchen geformt und dann in den Dünndarm weitergegeben. Im Dünndarm erweichen diese Kügelchen zu einer breiartigen Masse. Die Formung der Kotballen erfolgt später im Dickdarm.“
In den oberen Schichten des Magens können Diastase und Bakterien noch so lange wirksam sein, bis das Futter von der Magensalzsäure durchdrungen wird und die Reaktion von alkalisch nach sauer umschlägt.
Im sauren Bereich werden die von den Hauptzellen der Magenwand ausgeschiedenen Fermente Pepsin und Lab wirksam. Sie spalten Eiweiß zu Albumosen und Peptone, deren weiterer Abbau durch die eiweißspaltenden Fermente des Dünndarms erfolgt. Ferner sind auch lipolytische (fettspaltende) Fermente vorhanden.
Kaninchen können, wie Pferde, nicht erbrechen (Dorn, 1972). Wird der Magen mit leichtgärenden Futterstoffen (z. B. jungem Klee) oder quellenden (z. B. Trockenschnitzel) gefüllt und/oder werden unzureichende Mengen an Salzsäure produziert, so kommt es zu einer starken Belastung des Magens, in Extremfällen bis zur Ruptur (Reißen) der Magenwand.
Wer Kaninchen längere Zeit beobachtet, wird feststellen, dass sie sehr häufig kleine Futtermengen aufnehmen. Nach Werner (1969) wurden 70-80 Mahlzeiten innerhalb von 24 Std. beobachtet, von denen eine Mahlzeit teilweise nur 1-2 Min. dauerte. Die Tatsache häufiger Aufnahme kleiner Futterportionen kann durch eigene Beobachtungen bestätigt werden (vgl. Dorn, 1973).
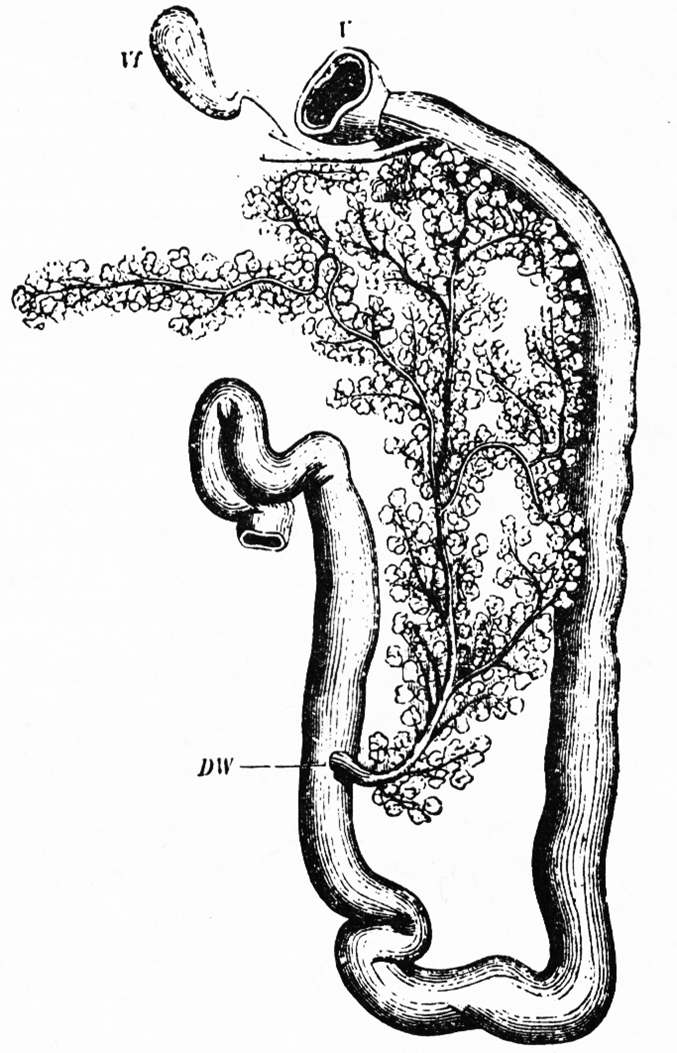
Abb. 8: Bauchspeicheldrüse und Gallenblase des Kaninchens mit Einmündung der Ausführungsgänge in die Schleife des Duodenums. V Pylorus, Vf Gallenblase (nach Krause aus Mangold, 1950).
Die Erhaltung der Magenfüllung durch fortlaufende Futteraufnahme bzw. die Schrumpfung des Magens bei Futterentzug sind offenbar für einen Weitertransport des Futters in den Dünndarm erforderlich. Dabei dürfte auch das Fressen des Blinddarmkotes (s. Kotfressen) eine Rolle spielen (vgl. Mangold, 1950). Die Reizung der Magendrüsen erfolgt durch Menge und Beschaffenheit der Futterstoffe, psychische Reize und zeitliche nach Gewöhnung. Bei längerer gleichmäßiger Ernährung stellt sich die Sekretion des Magens und auch des Darmes – auf die Zusammensetzung der Nahrung ein. Ein jäher Futterwechsel kann deshalb erhebliche Störungen hervorrufen, wenn die neue Nahrung zu ihrem Aufschluss eine andere Sekretzusammensetzung erfordert. Der Durchgang des Futters durch den niemals leeren Magen dauert nach Röntgenaufnahmen etwa 9 Stunden (Mangold, 1950).
Vorgänge im Dünndarm
Der z. T. verdaute oder für die Weiterverdauung vorbereitete Futterbrei gelangt vom Magen durch den Pförtner in den oberen Teil des Dünndarms, den Zwölffingerdarm. Hier mündet gleich unterhalb des Magenausganges der Gallengang und etwa 40 cm weiter der Ausgang der Bauchspeicheldrüse (Pankreas). Durch den alkalischen Bauchspeichel, der das eiweißspaltende Ferment Trypsin, ferner fett- und kohlehydratspaltende Fermente enthält, wird der saure Futterbrei neutralisiert. Die Gallensäuren bewirken eine feine Verteilung der Fette, die sich in der Form feinster Tröpfchen mit der Darmflüssigkeit mischen und eine Emulsion bilden. Dadurch wird der Abbau durch fettspaltende Fermente (Lipasen) erleichtert.
Die Bauchspeicheldrüse (Abb. 8) liefert nicht nur für die Verdauung wichtige Fermente, die in den Dünndarm fließen (DW), sondern produziert auch in den Langerhans’schen Inseln das für den Kohlehydratstoffwechsel und Blutzuckerspiegel wichtige Hormon Insulin. Bei Insulinmangel entsteht die Zuckerkrankheit (Diabetes mellitus), die beim Menschen weit verbreitet ist.
Darmeigene Drüsen liefern das für die weitere Eiweißverdauung erforderliche Erepsin sowie Fermente für den Abbau der Fette und Kohlehydrate.
Im Dünndarm erfolgt der wesentliche Teil der chemischen Verdauung. Eiweiße, Fette und Kohlehydrate werden durch die Wirkung der Fermente von der kolloidalen zur wasserlöslichen kristallinen Form abgebaut, in der sie in die Blutbahn gelangen können. Alle Fermente wirken spezifisch, d. h. sie sind jeweils nur auf einen bestimmten Nährstoff spezialisiert. Dabei wird Eiweiß über mehrere Stufen in die verschiedenen Aminosäuren, Kohlehydrate in Einfachzucker (Traubenzucker, Fruchtzucker, Laktose u. a.), Fette in Glyzerin und Fettsäuren zerlegt. Wasserunlösliche Fettsäuren werden durch Gallensäure und Alkali löslich gemacht.
Die Resorption (Aufsaugen) der verdauten Nährstoffe beginnt bereits im Magen. Der Hauptrest wird von der Darmschleimhaut aufgenommen. Der Dickdarm saugt den bisher noch nicht resorbierten Teil des Wassers auf. Bei Darmerkrankungen ist diese Resorption gestört, so dass dünnflüssiger Kot entleert wird.
Vorgänge im Dickdarm
Im Dickdarm (Abb. 1 u. 9) bieten besonders die beiden ersten geräumigen Abschnitte (Blinddarm und Colon) günstige Voraussetzungen für den bakteriellen Aufschluss der im Magen und Dünndarm noch nicht verdauten Nahrungsstoffe. Der noch erhebliche Wassergehalt des Futterbreies, eine im Wesentlichen neutrale bis schwachalkalische Reaktion, reichliche Bakteriennahrung und eine fortlaufende Durchmischung des Nahrungsbreies ermöglichen eine starke Vermehrung und Tätigkeit der Bakterienflora. Die wichtigsten Aufgaben einer physiologischen, d. h. dem Darm eines gesunden Kaninchens angepassten Darmflora sind: 1. Abbau der Cellulose (Rohfaser), dem Hauptbestandteil der pflanzlichen Zellwände, mit Hilfe des von den Cellulose-Bakterien erzeugten Fermentes Cellulase. 2. Eiweißveredelung und Eiweißsynthese Bakterien sind in der Lage, aus nicht vollwertigem pflanzlichen Eiweiß und nicht eiweißartigen Stickstoffverbindungen (z. B. Amiden) hochwertiges Bakterieneiweiß aufzubauen, das nach ihrem Tod den Wirtstieren zur Verfügung steht. Es herrscht hier also eine echte Symbiose, d. h. eine Zusammenarbeit zwischen den Bakterien und ihrem Wirt, dem Kaninchen, zu beiderseitigem Nutzen
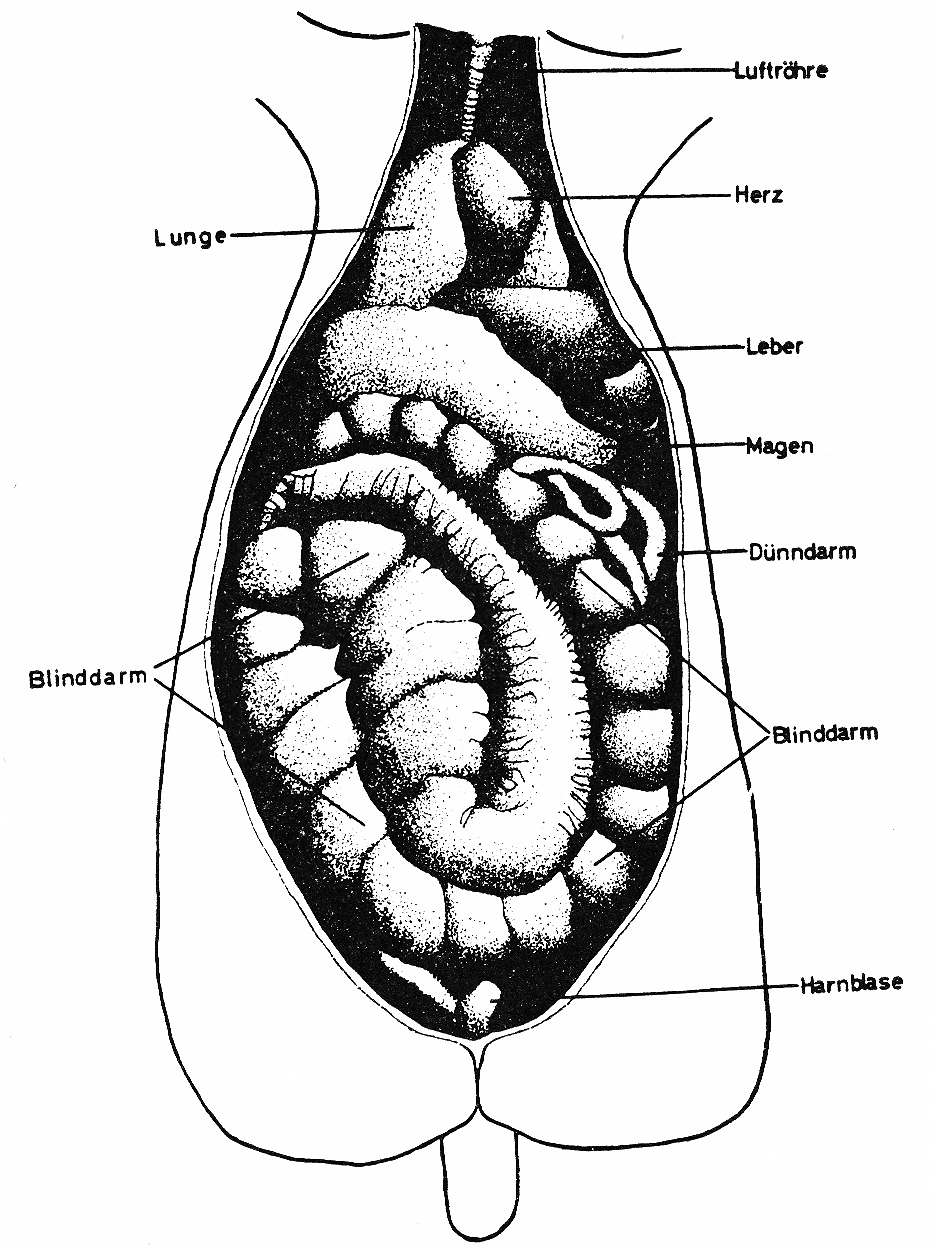
Abb. 9: Natürliche Lage der Körperorgane in einem erwachsenen Kaninchen. Rasse: Weiße Neuseeländer.
Diese Fähigkeit der Bakterien, Eiweiß auf- und umzubauen, ermöglicht es den Kaninchen und anderen Pflanzenfressern, mit biologisch nicht vollwertigem pflanzlichen Eiweiß auszukommen.
3. Vitaminsynthese Eine weitere nützliche Tätigkeit bestimmter Darmbakterien besteht in der Synthese (Aufbau) lebenswichtiger Vitamine der B-Gruppe und von Vitamin K2. Wenn diese Vitamine nicht bereits in ausreichenden Mengen im Futter enthalten sind, was bei der herkömmlichen Fütterung (Grünfutter, Saftfutter und Heu mit Zugaben von Getreidekörnern u. a.) oft nicht der Fall sein dürfte, ist diese Vitaminsynthese wichtig für das Leben und die Gesundheit der Tiere.
Hinsichtlich des Umfanges der Rohfaserverwertung beim Kaninchen bedürfen z. T. auch heute noch vorhandene Vorstellungen, nach denen Kaninchen in der Verwertung der Rohfaser mit den Wiederkäuern vergleichbar und zwischen Rind und Schwein einzuordnen seien, einer Korrektur. Wir wissen heute, dass man die Fähigkeit des Kaninchens auf diesem Gebiet überschätzt hat (vgl. Werner, 1969). Bei von Nehring (1955) veröffentlichten Fütterungsversuchen mit 14 Futterstoffen (Erdnüsse, Mais, Erdnuss-Extraktionsschrot, Sojabohnen, Erbsen, Gerste, Sojaextraktionsschrot, Erdnussexpeller, Leinsamen, Kokusextraktionsschrot, Hafer, Leinsamenextraktionsschrot, Palmkernexpeller und Palmkernextraktionsschrot) lag die Rohfaserverwertung beim Kaninchen mit 34,4 % hinter der von Rind, Schaf, Schwein und Ratte.
Die Verdauung der Rohfaser erfolgt sowohl im Blinddarm als auch im übrigen Teil des Dickdarms. Nach einer operativen Entfernung des Blinddarms ging die Rohfaserverwertung bei Versuchskaninchen auf etwa die Hälfte zurück (Zuntz und Ustjanzew, 1904/5).
Kotfressen
Eine früher für anomal und krankhaft gehaltene, seit langem aber als natürlich und für die Gesundheit der Tiere notwendig anerkannte Eigenart des Kaninchens ist das „Kotfressen", das auch beim Wildkaninchen und anderen Lagomorphen sowie Nagetieren, z. B. Ratten, Mäusen, Eichhörnchen u. a. beobachtet worden ist.
Nach den von Harder (1949 u. 1950) u. a. durchgeführten Untersuchungen wird ein spezieller, aus dem Blinddarm stammender weicher Kot, der über 30% des gesamten Kotes ausmacht, von den Kaninchen wieder aufgenommen. Dieser Blinddarmkot unterscheidet sich in seinem Aussehen und in seiner Zusammensetzung von den normalen Kotperlen (Abb. 10 u. Tab. 2). Blinddarmkot, deren einzelne Perlen einen Durchmesser von 3-11 mm aufweisen, wird im Gegensatz zu den normalen 10-12 mm großen, eirunden Perlen in langen, zusammengeklebten wurstähnlichen Gebilden abgesetzt. Diese haben eine mit Schleim überzogene glänzende Oberfläche. Die einzelnen weichen Perlen erhalten durch das Zusammen- drücken eine mehrkantige Form.
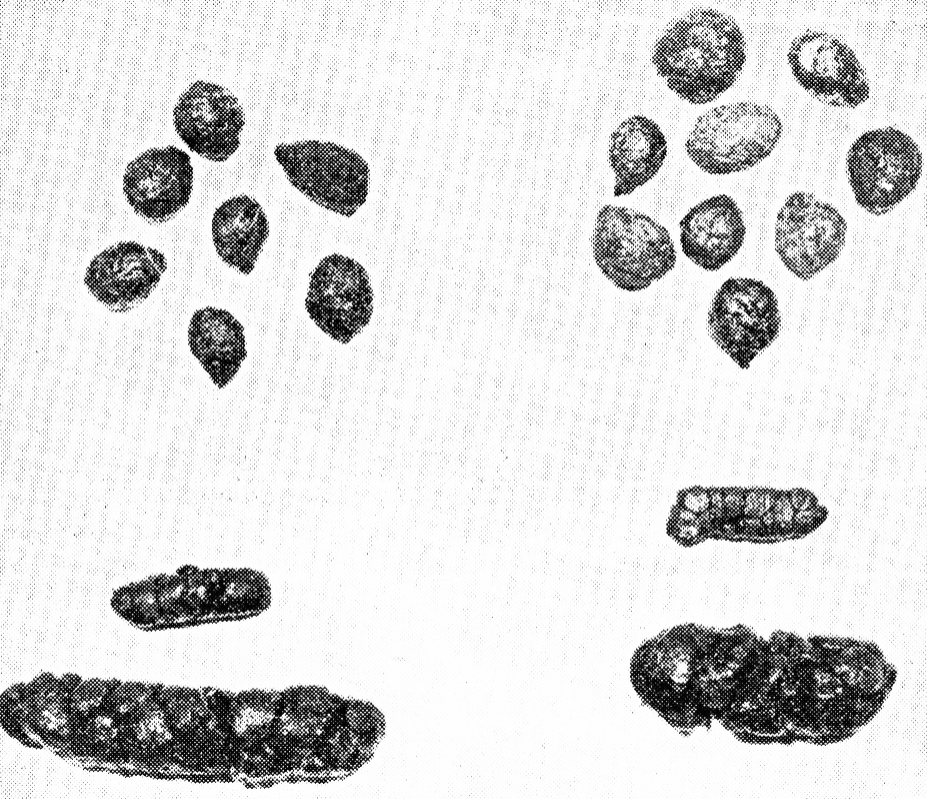
Abb. 10: Normale Kotperlen, oben, und Blinddarmkot, unten. Bei näherer Betrachtung erkennt man die aus vielen zusammengeklebten Kotperlen bestehende Blinddarmausscheidung.
Nach den Angaben von Blound (1957) findet die Formung dieser Blinddarm-Pellets ungefähr 6 Std. nach der letzten Mahlzeit, beim Hauskaninchen etwa gegen Mitternacht statt. Etwa 1 Std. später erscheint dieser Nasskot am Anus (After), von dem er direkt abgenommen und unzerkaut ab- geschluckt wird. Deshalb wird dieser Kot auch als Nachtkot be- zeichnet. Nach eigenen Untersuchungen scheiden Kaninchen auch am Tage weichen Kot aus, der mengenmäßig jedoch geringer ist als der Nachtkot. Wegen der direkten Abnahme des Blinddarmkotes vom After ist auch eine Versorgung der Tiere mit dieser Kotfraktion bei Haltung auf Gitterrosten gesichert. Gelegentlich findet man auch Blinddarmkot in der Streu oder auf den Kotblechen. Ferner wurde auch die Aufnahme normaler Kotperlen beobachtet. Der aufgenommene Blinddarmkot bleibt einige Stunden in der ursprünglichen Form im oberen Fundusteil des Magens erhalten. Danach wandert er unter Zersetzung zum Magenausgang. Dort wurden von uns bei geschlachteten Tieren niemals Kotperlen oder geformte Elemente gefunden. Tab. 2 zeigt die unterschiedliche Zusammensetzung von normalem Kot und Blinddarmkot.
Tab. 2
Analysen von normalem Kot und Blinddarmkot des Kaninchens (nach Hardeler, aus Harder, 1950)
Gehalte in % der Trockensubstanz

Wie die Zahlen zeigen, enthält der Blinddarmkot einen wesentlich höheren Wassergehalt und einen niedrigeren Fett- und Rohfasergehalt als der normale Kot, dagegen ist der Roheiweißgehalt 2,3mal, die Keimzahl 3,5mal so hoch wie im normalen Kot. Außerdem ist der in der Tab. 2 nicht angegebene Vitamingehalt des Blinddarmkotes um ein Mehrfaches höher als im normalen Kot. Die Coecotrophie hat vermutlich folgende physiologische Bedeutung:
1. Die von Blinddarmbakterien gebildeten Vitamine, die offenbar im Colon und Rektum nicht oder doch nicht in ausreichendem Maße resorbiert werden, bedürfen einer erneuten Aufnahme durch das Tier, um ihre Wirksamkeit entfalten zu können.
2. Auch der erhöhte Gehalt des Blinddarmkotes an hochwertigem Eiweiß und Spurenelementen wird von den Tieren durch eine erneute Aufnahme verwertet.
3. Es darf angenommen werden, dass die während der Nacht er- folgte Aufnahme des Blinddarmkotes durch Füllung des Magens zum Weitertransport des Futterbreies beiträgt. Kaninchen, die durch Anlegen eines Kragens an der Aufnahme des Blinddarmkotes gehindert wurden oder auf 105 Grad Celsius erhitzten Kot erhielten, zeigten eine erhöhte Anfälligkeit gegen Coccidiose und ein vermindertes Wachstum (Harder, 1950). Gesundheitliche Gefahren ergeben sich bei der Aufnahme von frischem Kot nicht, weil der Blinddarmkot normalerweise direkt vom After abgenommen wird und evtl. im Kot enthaltene Coccidienoocysten erst nach längerer Reifungszeit außerhalb des tierischen Organismus infektiös werden.
Jungtiere, die zunächst noch keinen Blinddarmkot bilden, können sich allerdings durch ein durch Kot verschmutztes Gesäuge der Häsin oder durch Aufnahme von 24-36 Std. gelagerten Häsinnenkot bzw. durch verschmutztes Futter mit Coccidien infizieren. Sauberkeit im Stall und trockene Stallböden sind wichtige Vorbeugungsmaßnahmen.
Beim Übergang des Futterbreies vom Dünndarm in den Dickdarm gelangt nur etwa 1/3 in den Blinddarm; etwa 2/3 werden direkt in den Grimmdarm (Colon) befördert.
Es bedarf weiterer Untersuchungen, ob dabei eine Sortierung des Futterbreies stattfindet, was nach der unterschiedlichen Zusammensetzung von Blinddarmkot und normalem Kot vermutet wird. Vielleicht spielt dabei der am Ende des Darmes befindliche Sacculus rotundus (Abb. 1) eine Rolle. Im Endabschnitt des Dickdarmes (Colon u. Rectum) wird die Flüssigkeit des Darminhaltes weitgehend resorbiert. Gleichzeitig beginnt die Formung der Kotballen, die durch peristaltische Darmbewegungen in Richtung After befördert und dort ausgeschieden werden.
Koprophagie oder Coecotrophie?
Der aus dem Griechischen stammende Fachausdruck für Kotfressen heißt „Koprophagie“. Da Kaninchen jedoch nicht die normalen Kotperlen, sondern das aus dem Blinddarm (Coecum) stammende vitamin- und eiweißreiche, für die Gesundheit und den normalen Stoffwechsel wichtige Produkt aufnehmen, das von Harder (1950) als „Blinddarmspeise“ bezeichnet wird, halte ich den Ausdruck Coecotrophie für diesen Vorgang besser geeignet als Koprophagie. Coecotrophie bedeutet, frei übersetzt: Ernährung durch den Blinddarmkot.
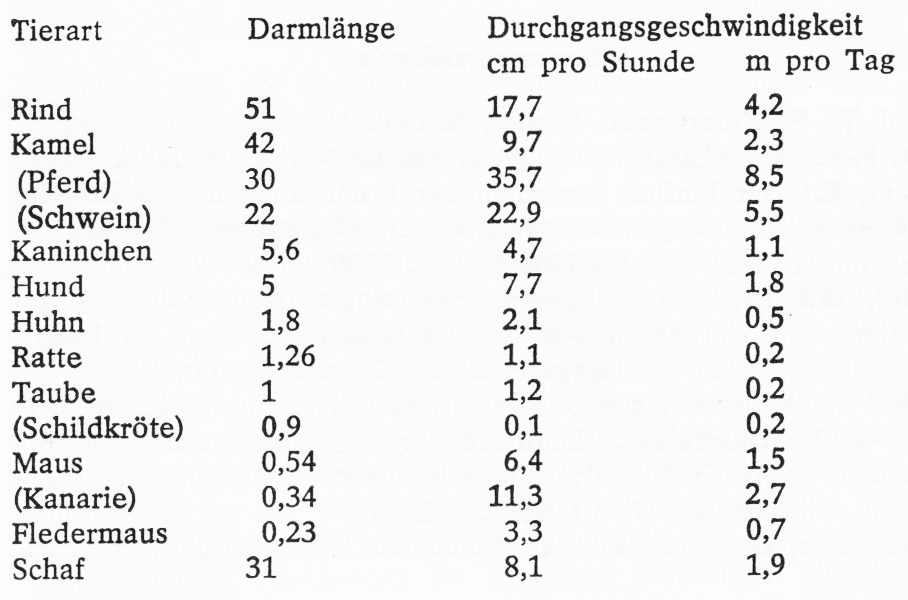
Durchgangszeiten und Durchgangsgeschwindigkeiten
Nach Mangold, Columbus und Hock (1937) begann die Entleerung eines Versuchsfutters bei Kaninchen zwischen 3,5 und 6,5, in der Regel bei 5 Std. nach der Aufnahme der Probemahlzeit. Normalerweise ist die Ausscheidung nach 5-6 Tagen beendet, wo- bei Art und Zusammensetzung der Nahrung einen beschleunigenden oder verzögernden Einfluss ausüben können.
Diese Dinge müssen z. B. bei Fütterungsversuchen berücksichtigt werden. Kenntnisse von diesen Vorgängen können aber auch für den Tierarzt und den Züchter wichtig sein. Interessant ist eine von Mangold (1951) gemachte Zusammenstellung über die Zusammenhänge von Darmlänge und Durchgangsgeschwindigkeit bei verschiedenen Tierarten.
Mangold weist auf die überraschend großen Unterschiede hin, die sich, wenn man von der Schildkröte als Kaltblüter absieht von 1,1 cm/Std. bei der Ratte bis 35,7 cm/Std. beim Pferd differieren. Beim Kaninchen wandert der Futterbrei 4,7 cm/Std., entsprechend 1,1 m täglich. Von einigen Überschneidungen abgesehen, kann man sagen, dass die Durchgangsgeschwindigkeit mit der Länge des Darmes abnimmt.
Tab. 3
Durchgangsgeschwindigkeit und absolute Darmlänge (geordnet nach den Darmlängen)
Schlussbemerkungen
Im vorstehenden Artikel ist versucht worden, einen Überblick über die Verdauungsvorgänge beim Kaninchen zu geben und dabei die gegenüber anderen Tierarten vorhandenen Besonderheiten der Funktion der Verdauungsorgane herauszustellen. Die Kenntnis der Zusammenhänge ist sowohl für den Praktiker als auch für den Wissenschaftler und Tierarzt bei der Fütterung der Tiere und der Durchführung von Versuchen und Behandlungsverfahren wichtig.
Viele Erkenntnisse konnten auf diesem Gebiet schon gesammelt werden. Weitaus mehr ist jedoch noch nicht oder doch nicht ausreichend bekannt, so dass eine weitere Klärung durch wissenschaftliche Versuche erforderlich ist.
Literaturverzeichnis
Blount, W. P.: Zitiert nach Werner, K. 1969 Dorn, F.-K.: Rassekaninchenzucht, Neumann Verlag, Radebeul, 1973
Haesler, K.: Der Einfluss verschiedener Ernährung auf die Größenverhältnisse des Magen-Darmkanals bei Säugetieren. Zeitschr. f. Tierzüchtung und Züchtungsbiologie, 17, 1930
Harder, W.: Zur Morphologie u. Physiologie des Blinddarms der Nagetiere. In: Verhandlungen der deutschen Zoologen in Mainz. K.-G. Leipzig, Akad. Verlagsgesellschaft, Geest & Portig, 1949
Harder: Mein Kaninchen frisst Kot. Dt. Kleintierzüchter, Nr. 14, 1950
Hoppe-Seyler/Thierfelder: Handbuch der polyzoologischen, pathologischen u. chemischen Analyse, herausgegeben von: Lange, K., Lehnartz, E., und Siebert, G., 10. Aufl., 5, 1953
Hochstrasser, G.: Wie kaut das Kaninchen? Dt. Kleintierzüchter 81, Nr. 22, 1972 Jung, S.: Grundlagen für die Zucht und Haltung der wichtigsten Versuchstiere. Gustav Fischer Verlag, Stuttgart, 1962
Kametaka, M.: Mikrobiologie der Verdauung beim Kaninchen. In: Biologie und Biochemie der mikrobiellen Verdauung. Giesecke u. Hendrickx, BLV Verlagsgesellschaft München, Bern, Wien, 1973
Lenkeit, W.: Zitiert nach Mangold, 1951
Mangold, E.: Die Verdauung bei den Nutztieren. Akad. Verlag, Berlin, 1950
Mangold: Darmlänge, Durchgangszeit und Durchgangsgeschwindigkeit. Akad. Verlag, Berlin, 1951
Mangold u. Haesler, K.: Der Einfluss verschiedener Ernährung bei Säugetieren (nach Versuchen an Ratten). Wissensch. Arch. f. Landw.. Abt. B: Tierernährung und Tierzucht, 2, 1930
Mangold, Columbus, A., Hock, A.: Arch. f. Geflügelk., 11, 1937 Nachtsheim, A.: Vom Wildtier zum Haustier. Verlag Paul Parey, Berlin, 2. Aufl., 1949 Nehring, K.: Lehrbuch der Tierernährung und Futtermittelkunde. Neumann Verlag, Radebeul u. Berlin 1952 u. 1955
Wetzel, G.: Anat. Anz. 71, Ergänzungsheft, 1931
Werner, K.: Kaninchenmast von Scheelje, Niehaus, Werner, Verlag Eugen Ulmer, Stuttgart, 1969
Zuntz, N., u. Ustjanzew, W.: Zitiert nach Magold, 1950 112


