

Professor Dr. Ruth Gruhn
Institut für Tierzucht und Haustiergenetik der Universität Göttingen
Direktor: Professor Dr. F. Haring
„Das Baue. Jahrbuch“ 1967
Bei keinem Haustier ist die Vererbung der Haarfarbe so gut analysiert wie bei dem Kaninchen, und es gibt auch keine Tierart, bei welcher ihre Züchter, weitgehend auf der Erfahrung aufbauend, nicht wissen, welche Fellfarbe sie bei der Kreuzung von zwei verschiedenfarbigen Rassen erwarten können. Da aber gelegentlich doch Unklarheiten bestehen, soll es die Aufgabe des vorliegenden Beitrages sein, kurz die genetischen Grundlagen der Haarfarbe und damit den Genotyp (Erbbild) der verschiedenen Rassen darzulegen, woraus sich zugleich die Aufspaltung bei Kreuzungen erklärt.
Einige grundsätzliche Betrachtungen über die Vererbung nach den Mendel'schen Regeln seien vorausgeschickt. Zur Darstellung der Genpaare, welche einer Merkmalsausprägung zugrunde liegen, verwendet man Buchstaben, z. B. A,a; B,b usf. Besitzen Individuen in dem einen Merkmal zugeordneten Genpaar die gleichen Gene (gleiche Allele) – das Erbbild oder der Genotyp ist also AA oder aa, BB oder bb – dann nennt man sie für dieses Genpaar reinerbig oder homozygot. Haben sie dagegen in diesem Genpaar verschiedene Allele, also Aa oder Bb, dann nennt man sie mischerbig oder heterozygot, gelegentlich auch spalterbig. Wenn man zwei sich in einem Genpaar unterscheidende reinerbige Eltern miteinander paart, dann ist die daraus hervorgehende Nachkommenschaft einheitlich. Hierbei sind aber zwei Fälle möglich:
1. Die Merkmalsausprägung der Nachkommen nimmt eine Zwischenstellung zwischen beiden Eltern ein. Man spricht in diesem Fall von intermediärer Vererbung. Ein Beispiel hierfür sind die vom Standard anerkannten Marderkaninchen (Genotyp aman), welche aus den dunklen Mardern (Genotyp amam) und den Russenkaninchen entstehen.
2. Die Nachkommen gleichen ausschließlich dem einen Elter. In diesem Fall bezeichnet man das der Merkmalsausprägung zugrundeliegende Gen als dominant, das nicht in Erscheinung tretende aber als rezessiv. Ein auch hier jedem Kaninchenzüchter bekanntes Beispiel ist die Haarlänge. Kreuzt man Kaninchen mit normallangem Haar (Genotyp VV) mit langhaarigen Kaninchen (Genotyp vv), dann hat die Nachkommenschaft normallanges Haar. Ihr Genotyp ist Vv.
Sowohl bei intermediärem als auch dominantem Erbgang ist aber die Nachkommenschaft heterozygot (s. Beispiel 1: aman, Beispiel 2: Vv). Die Paarung von in einem Anlagenpaar heterozygoten Tieren untereinander führt aber zu einer Nachkommenschaft, die in diesem Anlagenpaar eine Aufspaltung zeigt. Der Grund dafür liegt darin, dass ein solches Kaninchen zwei Sorten Keimzellen bilden kann, nämlich Keimzellen mit dem Gen am und solche mit der Erbanlage an im ersten Fall und entsprechend im zweiten Fall Keimzellen mit V und Keimzellen mit v. Jede Keimzelle hat zu ihrer Bildung die gleiche Wahrscheinlichkeit. Schematisch dargestellt ergeben sich bei der Paarung von in einem Anlagenpaar heterozygoten Individuen folgende Möglichkeiten für die Nachkommen:
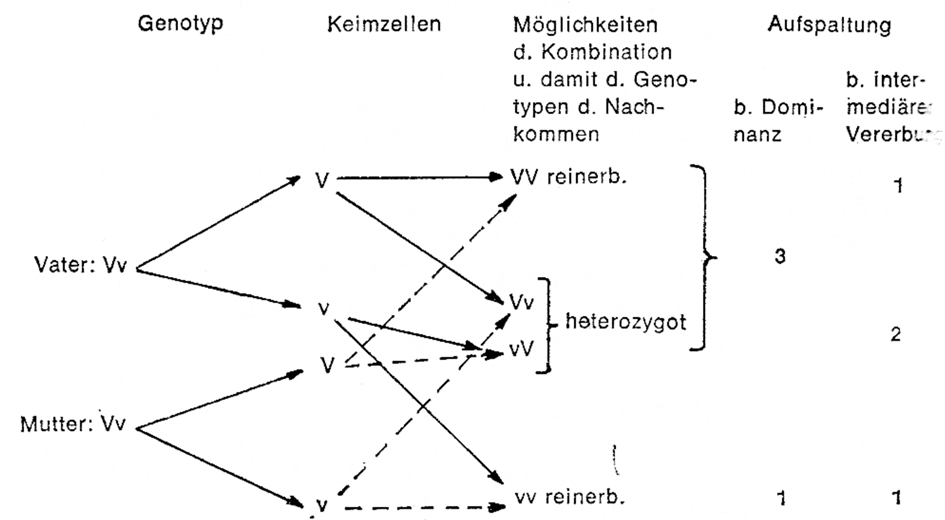
Jeder der vier Genotypen hat die gleiche Wahrscheinlichkeit, und es hängt vom Zufall ab, ob ein Kaninchen mit normal- langem oder langem Haar entsteht. Bei einer relativ großen Anzahl von Paarungen solcher heterozygoter Individuen wird man demzufolge bei dominanter Vererbung in der zweiten Nachkommengeneration eine Aufspaltung in 3 dominante Merkmalsträger (Genotyp VV Genotyp Vv in der Ausprägung): rezessiv (vv) erhalten. Bei intermediärer Vererbung aber liegen die heterozygoten Genotypen in ihrer Merkmalsausprägung zwischen den beiden Rassen und demzufolge kann man hier eine Aufspaltung 1:2:1 erwarten.
Diesen beiden Grundformen der Vererbung folgen ganz allgemein gesagt die der Haarfärbung zugrunde liegenden Erbanlagen:
Die Haarfarbe der Kaninchen wird im Wesentlichen durch 5 Anlagenpaare gesteuert, welche im Deutschen mit den Buchstaben A, B, C, D, G bezeichnet werden. Dabei bewirkt das Vorhandensein oder Fehlen dieser Faktoren jeweils eine andere Farbtönung. Im Einzelnen steuern diese Erbanlagen folgende Ausprägungen:
1. Anlagenpaar: A Pigmentgrundfaktor
(A muss vorhanden sein, damit überhaupt Pigment gebildet werden kann)
a Fehlen des Pigmentfaktors, weiße Farbe
2. Anlagenpaar: G Wildfarbfaktor oder Agoutifaktor
Er bewirkt die zonenweise Anordnung des Pigmentes (schwarz, blau, braun, gelb) im einzelnen Haar, welche zusammenwirkend einen graubraunen Farbton (Wildfarbe) ergibt.
g Fehlen des Wildfarbfaktors; die zonenweise Anordnung des Pigmentes im einzelnen Haar unterbleibt, das Kaninchen ist einfarbig gefärbt.
3. Anlagenpaar: B Farbfaktor Schwarz
b Farbfaktor Gelb
4. Anlagenpaar: C Farbfaktor Schwarz
c Farbfaktor Braun
5. Anlagenpaar: D Farbfaktor Schwarz
d Farbfaktor Blau
Das Wildkaninchen hat den Genotyp AABBCCDDGG. Durch plötzlich auftretende Mutationen („Erbänderung“), wie sie im Laufe der geschichtlichen Entwicklung bei jeder Tierart immer wieder festgestellt werden, kann jedes der oben angeführten Gene eine Zustandsänderung erfahren, welche durch sein Allel symbolisiert wird, d. h. es kann z. B. das Gen G in sein Allel g abgeändert werden. In einem solchen Fall unterbleibt die für das Wildkaninchen typische zonenweise Anordnung des Pigmentes, das dunklere Pigment deckt die helleren zu und das Haar erscheint völlig schwarz. Fehlt außerdem einer der Farbfaktoren für schwarzes Pigment (B, C oder D), dann tritt eine Farbverdünnung ein und je nachdem, welche der Farbfaktoren B, C, D fehlen, erhält man eine schwächere oder stärkere Aufhellung; hierbei kann jeder der möglichen Genotypen den Wildfarbfaktor G führen oder auch nicht. Nachfolgend sind die auf den oben genannten Genpaaren beruhenden Kombinationen mit ihrer Ausprägung in der Fellfarbe zusammengestellt. Dabei soll aus technischen Gründen und zur Vereinfachung der Übersicht von jedem Genpaar nur eines aufgezeichnet werden. Selbstverständlich führt jedes farbige Kaninchen und damit jeder Genotyp das Gen A; denn sein Vorhandensein ist Voraussetzung dafür, dass überhaupt Farbe gebildet werden kann.
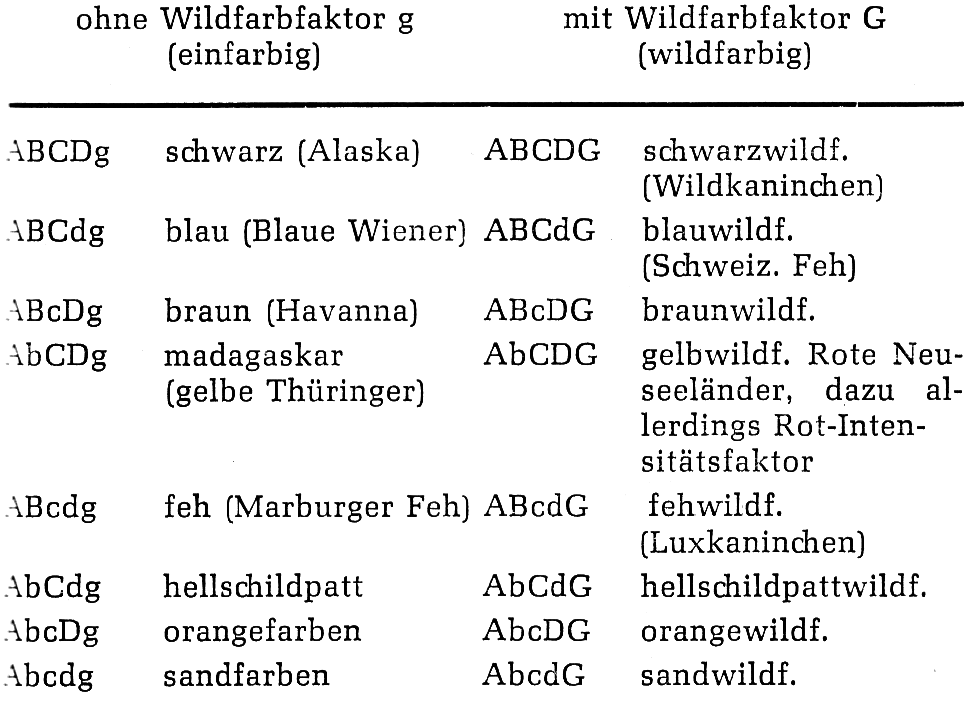
Da innerhalb der einzelnen Anlagenpaare Dominanz besteht, ist es stets möglich, dass ein Kaninchen in einem der Anlagenpaare heterozygot ist, ohne dass dieses äußerlich in Erscheinung tritt. Diese Heterozygotie wird erst dann erkannt, wenn zwei im gleichen Anlagenpaar heterozygote Tiere miteinander gepaart werden; denn dann kann der in diesem Anlagenpaar rezessive Genotyp ausspalten. Werden z. B. zwei in dem Anlagenpaar D heterozygote schwarze Kaninchen gepaart, dann können blaue Kaninchen ausspalten:
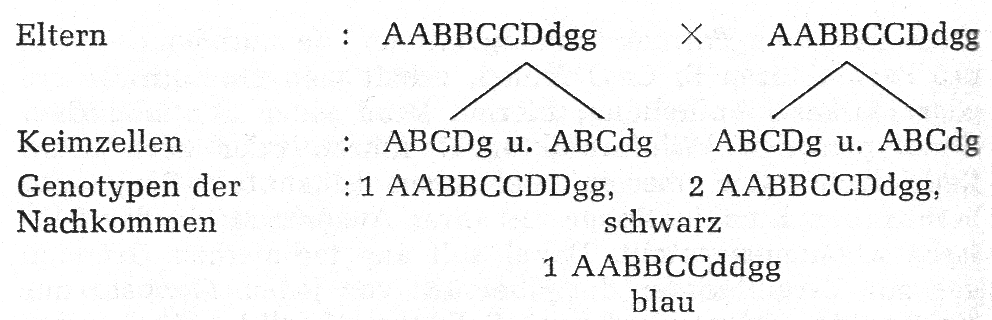
Würden die Alaska z. B. in dem Anlagenpaar B heterozygot sein (Bb), dann kann hier entsprechend der Genotyp AAbb- CCDDgg madagaskarfarben ausspalten.
Gleichzeitig ist aber auch erkennbar, dass man „Blau" nicht durch Kreuzung mit schwarzen Kaninchen intensivieren kann, wie es gelegentlich noch angenommen wird. Die Paarung von schwarzen Kaninchen mit blauen gibt bei Reinerbigkeit der schwarzen nur schwarze Nachkommen, bei Mischerbigkeit der schwarzen, aber schwarze oder blaue:
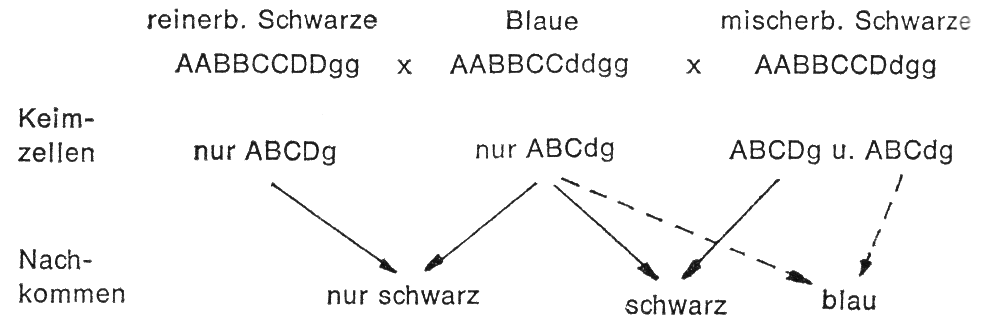
Aus der obigen Übersicht geht weiterhin hervor, dass z. B. die Paarung von Blauen Wienern mit Havanna auf Grund der Dominanz innerhalb der Anlagenpaare nur schwarze Nachkommen bringt.
Selbstverständlich kann auch der Fall vorkommen, dass an Stelle des Gens A sein rezessives Allel a tritt. Dann kann, wie oben schon dargelegt ist, auch bei Vorhandensein der Farbfaktoren B, C, D kein Pigment gebildet werden, und man erhält einen Albino. Albinos sind für die Anlage a immer reinerbig, also stets aa-Typen. Welche Farbfaktoren ein Albino außerdem führt (z. B. aaBBCCDDGG oder aabbcc- Ddgg usf.), das ist ihm äußerlich nicht anzusehen und wird nur in der Aufspaltung bei Anpaarung an farbige Kaninchen erkennbar.
Von dem Albino-Weiß zu unterscheiden ist noch das Weiß der Weißen Wiener, welche blaue Augen haben. Dieses Weiß beruht auf einem weiteren Anlagenpaar xx, wobei der Wild- typ das dominante Gen XX führt. Dieses rezessive x-Gen besitzt die Fähigkeit, die Haar-Farb-Ausbildung aller Farbfaktoren zu unterdrücken, so dass hier nur das Auge normal gefärbt ist. Ebenso wie alle farbigen Kaninchen führt auch das Albino das dominante Gen X, so dass sich Albinos und weiße Wiener in ihren Erbformeln in folgender Form unterscheiden:
Albinos : aa… XX (… = verschied. Faktoren)
Weiße Wiener : AA… XX (… = verschied. Faktoren)
Daraus geht zugleich hervor, dass die Paarung von Albinos mit Weißen Wienern eine Nachkommenschaft ergibt, die je nach den vorhandenen Farbfaktoren gefärbt ist; denn diese Nachkommenschaft ist in den Anlagenpaaren A und X heterozygot (Aa…Xx), d. h. es kann Farbe gebildet werden (Gen A) und das Farbunterdrückungsgen x ist rezessiv zu X.
Aus der Zusammenstellung der verschiedenen Genotypen und der Rassen, die diese repräsentieren (s. oben), geht hervor, dass noch längst nicht alle verschiedenfarbigen Rassen erfasst sind. In der Tat hat man es auch bei den aufgezeigten Anlagenpaaren in einzelnen Fällen nicht mit einem Anlagen- paar (A,a) zu tun, sondern zu dem Gen A gibt es mehrere Allele, die jeweils miteinander kombiniert auftreten können Die Ursache dafür liegt darin, dass das Gen A sich bei der Erbänderung nicht sofort in den Zustand des Gens a umgewandelt hat, sondern es erfolgte diese Erbänderung stufen weise:
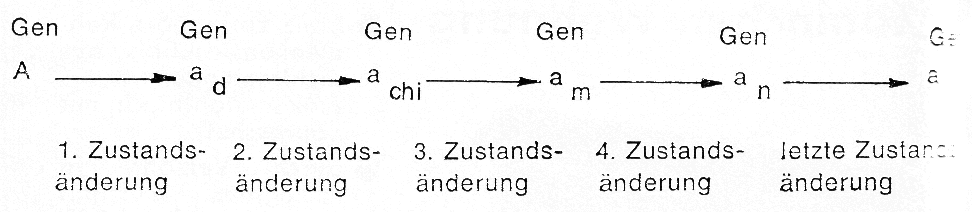
Die erste Zustandsänderung führt noch zu einer fast voll ständigen Ausfärbung, dem Dunkelchinchilla. Die zweite Zustandsänderung führt zu dem in der Kaninchengenetik mi achiachi bezeichneten Gen, weil dieser Farbfaktor die Chinchillafarbe bedingt. Noch weiter abgebaut als beim Chinchilla ist das Pigment beim Marderkaninchen, und dieses verkörpern auch die nächste Stufe der Erbänderung des Gens A, es wird mit am bezeichnet. Noch weniger Pigment gebildet wird bei der weiteren Zustandsänderung des Allels am in das Alle an, den sogenannten Russenfaktor, und erst diesem schließ sich als letzte Zustandsänderung das Allel a an. Das besagt an dem gleichen Genort der einander entsprechenden Chromosomen können die Gene A, adachi, am, an, oder a liegen, wobei jeweils die erstgenannten Gene gegenüber den nachfolgenden dominant sind. Man nennt diese Serie von Allelen auch die „Albinoserie" und Kaninchen, die der Farbe nach sein.

Die Farbe der „Russen"-Abzeichen (Schwarz-, Blau-, Gelb) richtet sich hierbei nach den vorhandenen Farbfaktoren. Ähnlich wie für das Gen A, so liegt auch für das Gen B eine solche Serie von Allelen vor, wobei hier nicht B das Anfangsglied ist, sondern ein mit dem Symbol Be bezeichnetes Gen. Dieses bedingt stärkste Pigmentierung. Reinerbige BeBe-Tiere sind fast schwarz, BeB-Tiere aber eisengrau. Die Erbformel der eisengrauen Deutschen Widder ist mithin AABEBCCDDGG. Der Erbänderung von B in das Allel b ist noch zwischengeschaltet die mit dem Symbol bj bezeichnete Zustandsform, welche sich in einer schwarz-gelben Bänderung der Japanerzeichnung ausprägt (Erbformel der Japaner AAbjbjCCDDGG). Diese Gelbserie umfaßt somit in der Reihenfolge der Dominanz:

Auch dem Wildfarbfaktor G steht nicht nur das Allel gegenüber, sondern hier ist ebenfalls ein Allel go zwischengeschaltet, welches die Lohzeichnung bedingt; diese Serie ist mithin
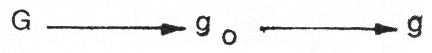
wobei G dominant zu go und g, go dominant zu g ist. Ein Kaninchen der Erbformel AABBCCDDgogo repräsentiert ein schwarz-gelbes Lohkaninchen und die für das Schwarzloh- Kaninchen typische rötliche Lohzeichnung wird durch das Hin- zutreten von Rot-Intensitätsfaktoren y1, y2… (beim Hasenkaninchen vorkommend) bewirkt. Ein Blaulohkaninchen würde dementsprechend die Erbformel AABBCCddgogo haben. Selbstverständlich ist Mischerbigkeit in den dominant vor- kommenden Anlagenpaaren (z. B. A, B, C) niemals auszuschließen, aber bei über viele Jahre reingezogenen Rassen unwahrscheinlich. Dieses gilt nicht nur für das hier zuletzt betrachtete Beispiel der Lohkaninchen, sondern auch für jede andere Rasse. Anhand der Erbformel, welche die Farbausprägung jeder Rasse bestimmt, kann sich jeder Züchter leicht ein Bild davon machen, welche Haarfärbung er bei der Kreuzung von zwei verschiedenen reingezüchteten Rassen erwarten kann. Hierzu ist letzthin nur das weiter oben dargelegte Wissen um das Zusammenwirken der Farbfaktoren B, C, D mit ihren Allelen notwendig und die Überlegung, welche Art von Keimzellen jede Rasse bilden kann; denn aus der Vereinigung der männlichen und weiblichen Keimzellen mit ihren jeweiligen Farbfaktoren erwächst das neue Individuum, das den aus der Kombination der väterlichen und mütterlichen Farbfaktoren resultierenden Genotyp repräsentiert.




